

mTOR signaling in VIP neurons regulates circadian clock synchrony and olfaction
- PMID: 29555746
- PMCID: PMC5889665
- DOI: 10.1073/pnas.1721578115
mTOR signaling in VIP neurons regulates circadian clock synchrony and olfaction
Abstract
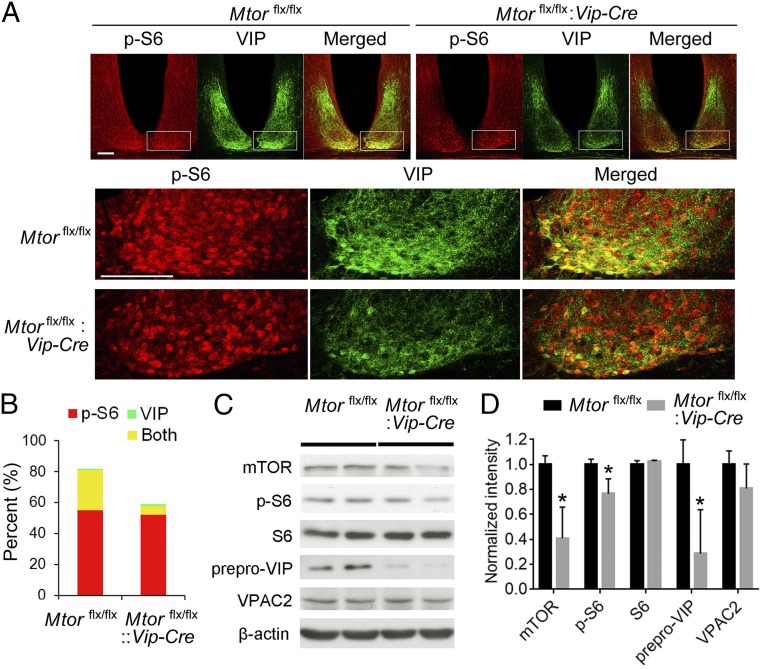
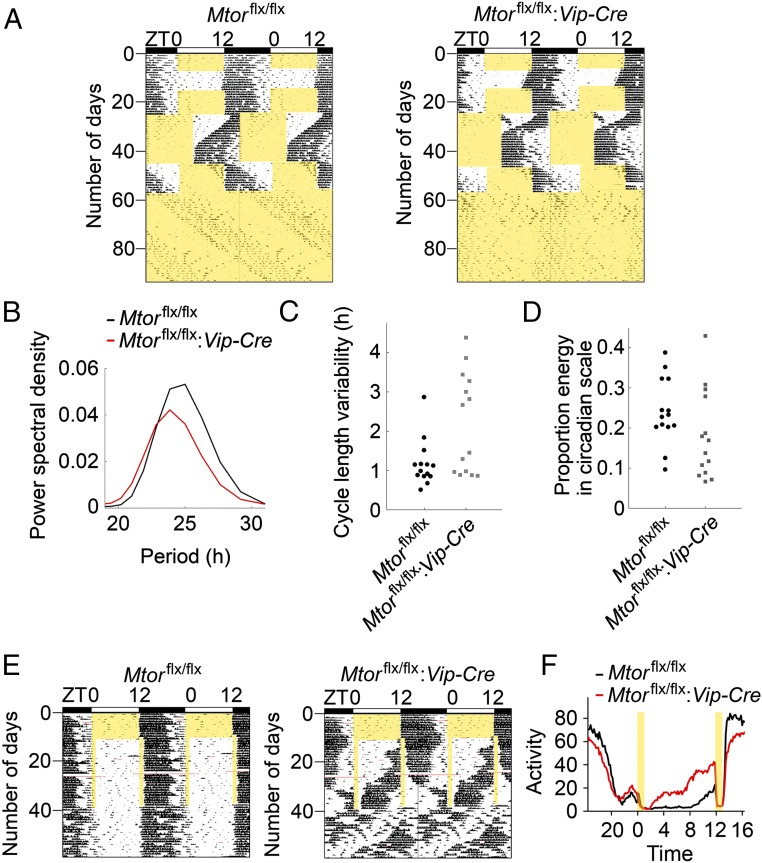
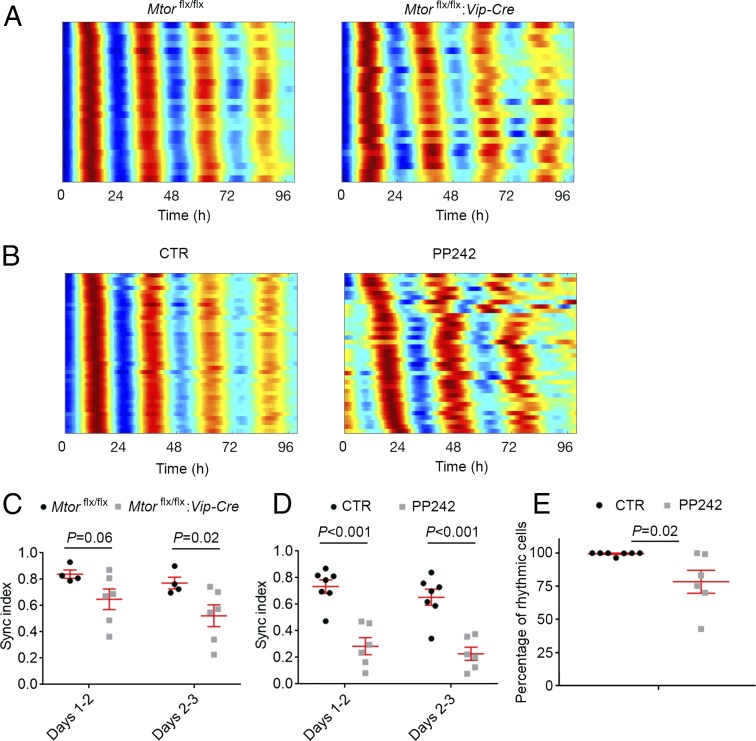
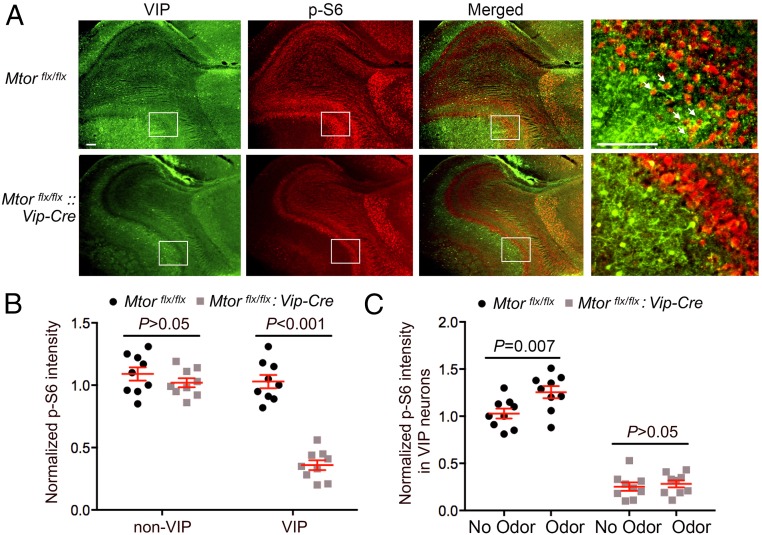
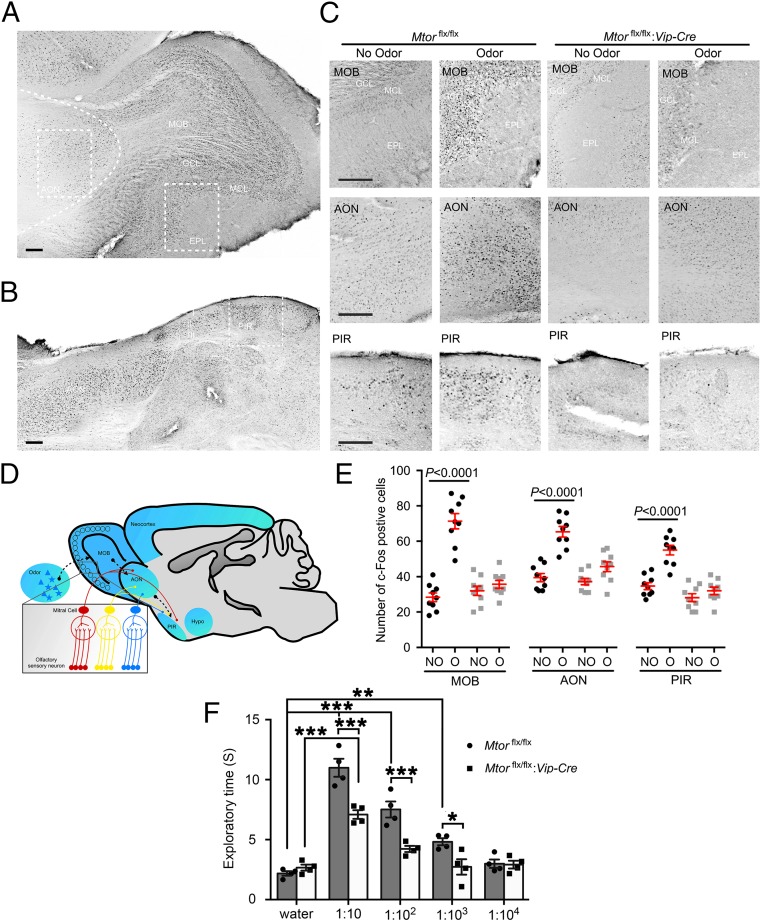
Mammalian/mechanistic target of rapamycin (mTOR) signaling controls cell growth, proliferation, and metabolism in dividing cells. Less is known regarding its function in postmitotic neurons in the adult brain. Here we created a conditional mTOR knockout mouse model to address this question. Using the Cre-LoxP system, the mTOR gene was specifically knocked out in cells expressing Vip (vasoactive intestinal peptide), which represent a major population of interneurons widely distributed in the neocortex, suprachiasmatic nucleus (SCN), olfactory bulb (OB), and other brain regions. Using a combination of biochemical, behavioral, and imaging approaches, we found that mice lacking mTOR in VIP neurons displayed erratic circadian behavior and weakened synchronization among cells in the SCN, the master circadian pacemaker in mammals. Furthermore, we have discovered a critical role for mTOR signaling in mediating olfaction. Odor stimulated mTOR activation in the OB, anterior olfactory nucleus, as well as piriform cortex. Odor-evoked c-Fos responses along the olfactory pathway were abolished in mice lacking mTOR in VIP neurons, which is consistent with reduced olfactory sensitivity in these animals. Together, these results demonstrate that mTOR is a key regulator of SCN circadian clock synchrony and olfaction.
Keywords: SCN; VIP; circadian clock; mTOR; olfaction.
Copyright © 2018 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Translational control of entrainment and synchrony of the suprachiasmatic circadian clock by mTOR/4E-BP1 signaling.Neuron. 2013 Aug 21;79(4):712-24. doi: 10.1016/j.neuron.2013.06.026. Neuron. 2013. PMID: 23972597 Free PMC article.
-
A circadian clock in the olfactory bulb controls olfactory responsivity.J Neurosci. 2006 Nov 22;26(47):12219-25. doi: 10.1523/JNEUROSCI.3445-06.2006. J Neurosci. 2006. PMID: 17122046 Free PMC article.
-
Vasoactive intestinal polypeptide mediates circadian rhythmicity and synchrony in mammalian clock neurons.Nat Neurosci. 2005 Apr;8(4):476-83. doi: 10.1038/nn1419. Epub 2005 Mar 6. Nat Neurosci. 2005. PMID: 15750589 Free PMC article.
-
An essential role for peptidergic signalling in the control of circadian rhythms in the suprachiasmatic nuclei.J Neuroendocrinol. 2003 Apr;15(4):335-8. doi: 10.1046/j.1365-2826.2003.01005.x. J Neuroendocrinol. 2003. PMID: 12622830 Review.
-
Neuroglial and synaptic rearrangements associated with photic entrainment of the circadian clock in the suprachiasmatic nucleus.Eur J Neurosci. 2010 Dec;32(12):2133-42. doi: 10.1111/j.1460-9568.2010.07520.x. Eur J Neurosci. 2010. PMID: 21143667 Review.
Cited by
-
Control of Selective mRNA Translation in Neuronal Subcellular Compartments in Health and Disease.J Neurosci. 2023 Nov 1;43(44):7247-7263. doi: 10.1523/JNEUROSCI.2240-22.2023. J Neurosci. 2023. PMID: 37914402 Free PMC article. Review.
-
A role for translational regulation by S6 kinase and a downstream target in inflammatory pain.Br J Pharmacol. 2021 Dec;178(23):4675-4690. doi: 10.1111/bph.15646. Epub 2021 Sep 12. Br J Pharmacol. 2021. PMID: 34355805 Free PMC article.
-
The Case for the Target of Rapamycin Pathway as a Candidate Circadian Oscillator.Int J Mol Sci. 2023 Aug 27;24(17):13307. doi: 10.3390/ijms241713307. Int J Mol Sci. 2023. PMID: 37686112 Free PMC article. Review.
-
Clock at the Core of Cancer Development.Biology (Basel). 2021 Feb 14;10(2):150. doi: 10.3390/biology10020150. Biology (Basel). 2021. PMID: 33672910 Free PMC article. Review.
-
Somatic mosaicism and interneuron involvement in mTORopathies.Trends Neurosci. 2025 May;48(5):362-376. doi: 10.1016/j.tins.2025.02.009. Epub 2025 Mar 22. Trends Neurosci. 2025. PMID: 40121168 Review.
References
-
- Wullschleger S, Loewith R, Hall MN. TOR signaling in growth and metabolism. Cell. 2006;124:471–484. - PubMed
-
- Ferrari S, Bandi HR, Hofsteenge J, Bussian BM, Thomas G. Mitogen-activated 70K S6 kinase. Identification of in vitro 40 S ribosomal S6 phosphorylation sites. J Biol Chem. 1991;266:22770–22775. - PubMed
-
- Weng QP, et al. Regulation of the p70 S6 kinase by phosphorylation in vivo. Analysis using site-specific anti-phosphopeptide antibodies. J Biol Chem. 1998;273:16621–16629. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
