

Mechanisms underlying the lack of endogenous processing and CLIP-mediated binding of the invariant chain by HLA-DP84Gly
- PMID: 29555965
- PMCID: PMC5859192
- DOI: 10.1038/s41598-018-22931-4
Mechanisms underlying the lack of endogenous processing and CLIP-mediated binding of the invariant chain by HLA-DP84Gly
Abstract
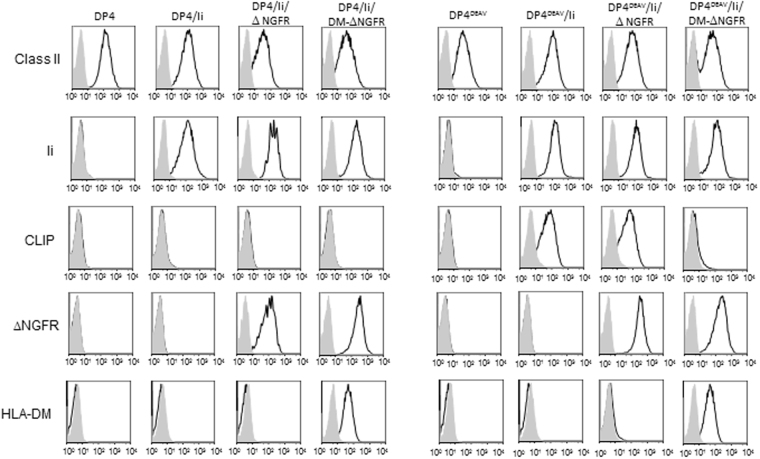
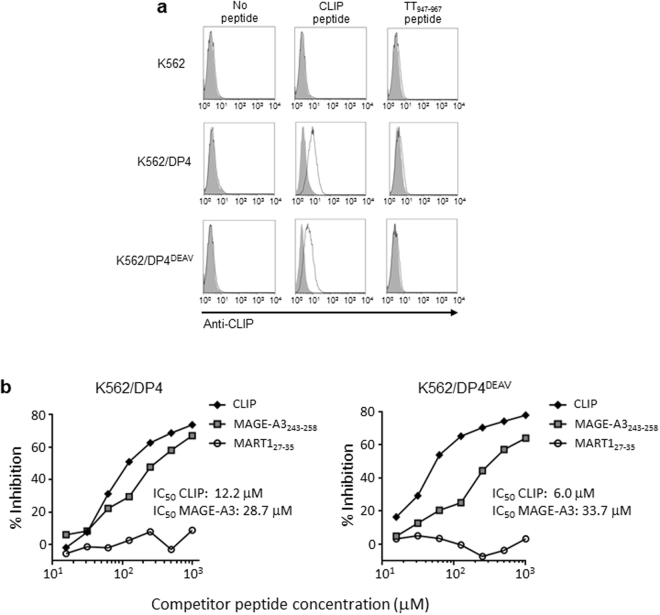
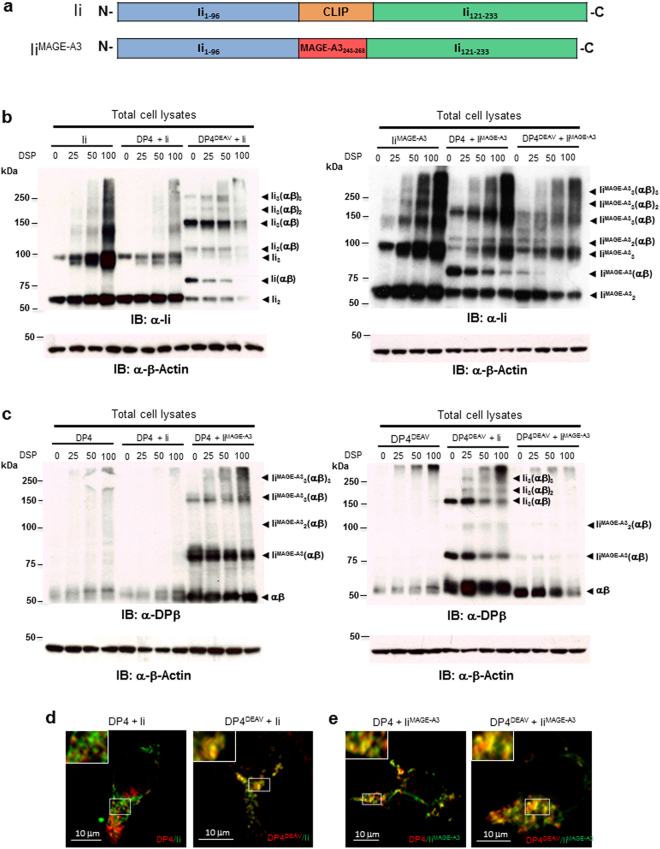
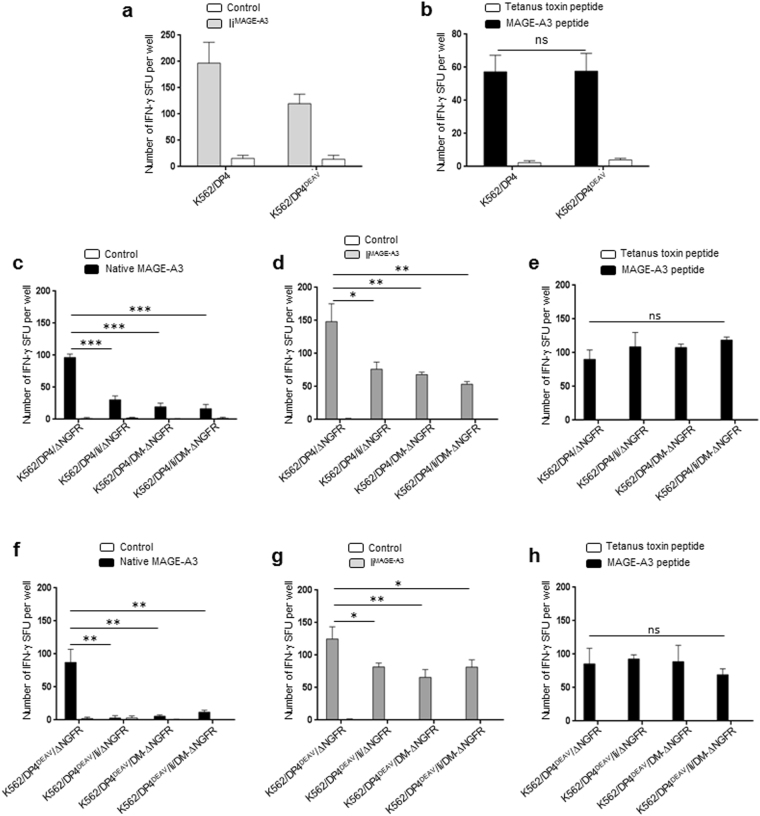
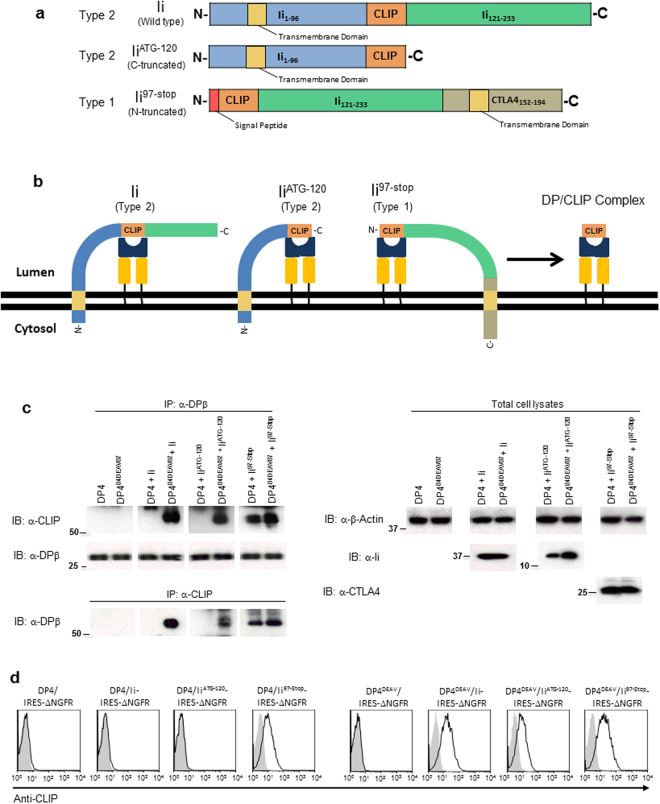
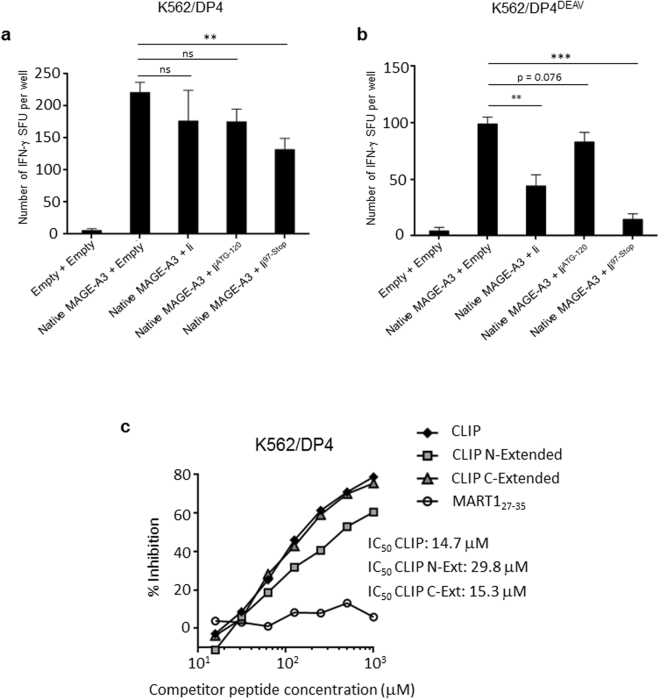
While the principles of classical antigen presentation via MHC class II are well-established, the mechanisms for the many routes of cross-presentation by which endogenous antigens become associated with class II molecules are not fully understood. We have recently demonstrated that the single amino acid polymorphism HLA-DPβ84Gly (DP84Gly) is critical to abrogate class II invariant chain associated peptide (CLIP) region-mediated binding of invariant chain (Ii) to DP, allowing endoplasmic reticulum (ER)-resident endogenous antigens to constitutively associate with DP84Gly such as DP4. In this study, we demonstrate that both the CLIP and N-terminal non-CLIP Ii regions cooperatively generate an Ii conformation that cannot associate with DP84Gly via the CLIP region. We also demonstrate the ability of DP4 to efficiently process and present antigens encoded in place of CLIP in a chimeric Ii, regardless of wild type Ii and HLA-DM expression. These data highlight the complex interplay between DP polymorphisms and the multiple Ii regions that cooperatively regulate this association, ultimately controlling the presentation of endogenous antigens on DP molecules. These results may also offer a mechanistic explanation for recent studies identifying the differential effects between DP84Gly and DP84Asp as clinically relevant in human disease.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
HLA-DP84Gly constitutively presents endogenous peptides generated by the class I antigen processing pathway.Nat Commun. 2017 May 10;8:15244. doi: 10.1038/ncomms15244. Nat Commun. 2017. PMID: 28489076 Free PMC article.
-
Efficient presentation of known HLA class II-restricted MAGE-A3 epitopes by dendritic cells electroporated with messenger RNA encoding an invariant chain with genetic exchange of class II-associated invariant chain peptide.Cancer Res. 2003 Sep 1;63(17):5587-94. Cancer Res. 2003. PMID: 14500399
-
Conformational alterations during biosynthesis of HLA-DR3 molecules controlled by invariant chain and HLA-DM.Eur J Immunol. 2001 Apr;31(4):1029-36. doi: 10.1002/1521-4141(200104)31:4<1029::aid-immu1029>3.0.co;2-q. Eur J Immunol. 2001. PMID: 11298327
-
What to do with HLA-DO/H-2O two decades later?Immunogenetics. 2019 Mar;71(3):189-196. doi: 10.1007/s00251-018-01097-3. Epub 2019 Jan 26. Immunogenetics. 2019. PMID: 30683973 Free PMC article. Review.
-
Achieving stability through editing and chaperoning: regulation of MHC class II peptide binding and expression.Immunol Rev. 2005 Oct;207:242-60. doi: 10.1111/j.0105-2896.2005.00306.x. Immunol Rev. 2005. PMID: 16181341 Review.
Cited by
-
Regulation of NK-Cell Function by HLA Class II.Front Cell Infect Microbiol. 2020 Feb 18;10:55. doi: 10.3389/fcimb.2020.00055. eCollection 2020. Front Cell Infect Microbiol. 2020. PMID: 32133304 Free PMC article.
-
Multiple autoimmunity and epitope spreading in monozygotic twins.J Transl Autoimmun. 2021 Nov 6;4:100132. doi: 10.1016/j.jtauto.2021.100132. eCollection 2021. J Transl Autoimmun. 2021. PMID: 34816112 Free PMC article.
-
Affinity-matured HLA class II dimers for robust staining of antigen-specific CD4+ T cells.Nat Biotechnol. 2021 Aug;39(8):958-967. doi: 10.1038/s41587-021-00836-4. Epub 2021 Mar 1. Nat Biotechnol. 2021. PMID: 33649568
-
Comprehensive Analysis of CD4+ T Cell Responses to CMV pp65 Antigen Restricted by Single HLA-DR, -DQ, and -DP Allotype Within an Individual.Front Immunol. 2021 Feb 15;11:602014. doi: 10.3389/fimmu.2020.602014. eCollection 2020. Front Immunol. 2021. PMID: 33658991 Free PMC article.
-
HLA associations in inflammatory arthritis: emerging mechanisms and clinical implications.Nat Rev Rheumatol. 2019 Jun;15(6):364-381. doi: 10.1038/s41584-019-0219-5. Nat Rev Rheumatol. 2019. PMID: 31092910 Review.
References
-
- Wilson NA, et al. Journal of immunology (Baltimore, Md.: 1950) 1998. Invariant chain can bind MHC class II at a site other than the peptide binding groove; pp. 4777–4784. - PubMed
-
- Layet C, Germain RN. Invariant chain promotes egress of poorly expressed, haplotype-mismatched class II major histocompatibility complex A alpha A beta dimers from the endoplasmic reticulum/cis-Golgi compartment. Proceedings of the National Academy of Sciences of the United States of America. 1991;88:2346–2350. doi: 10.1073/pnas.88.6.2346. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
