

Molecular mechanisms driving transcriptional stress responses
- PMID: 29556092
- PMCID: PMC6036639
- DOI: 10.1038/s41576-018-0001-6
Molecular mechanisms driving transcriptional stress responses
Abstract
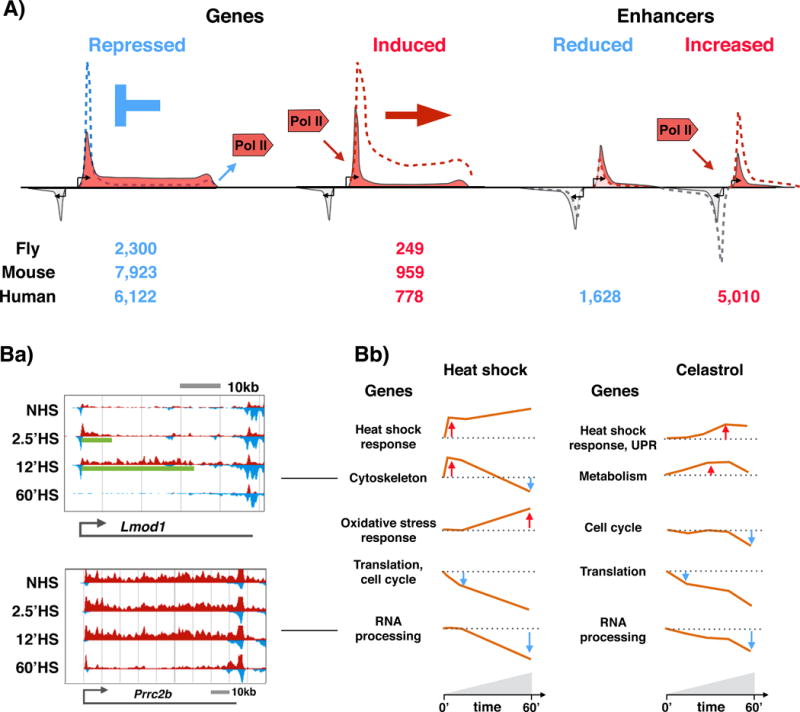
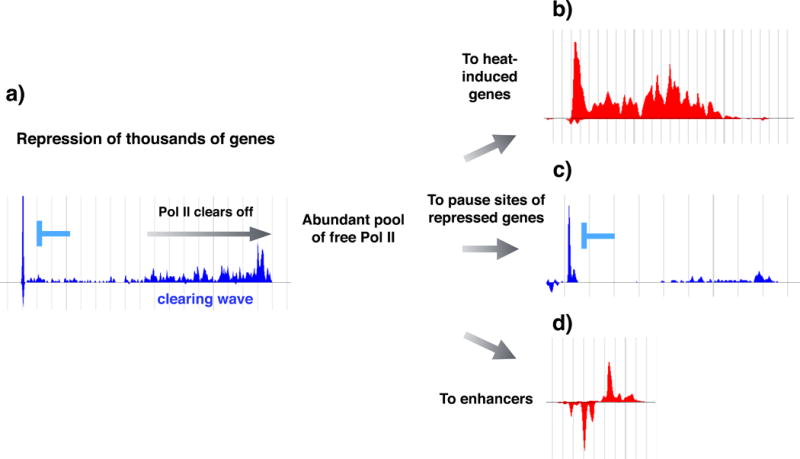
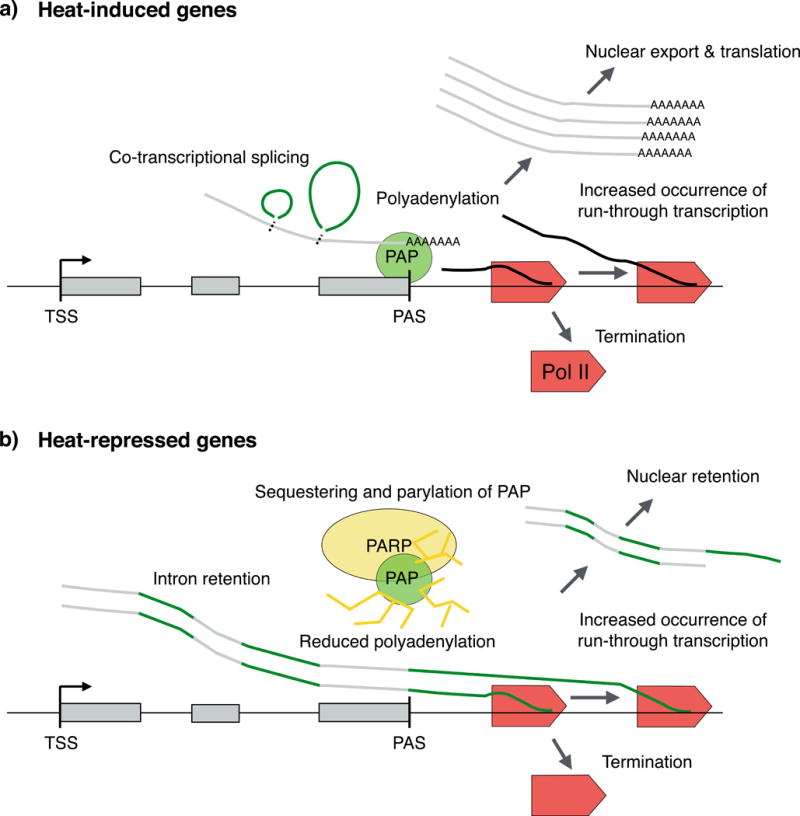
Proteotoxic stress, that is, stress caused by protein misfolding and aggregation, triggers the rapid and global reprogramming of transcription at genes and enhancers. Genome-wide assays that track transcriptionally engaged RNA polymerase II (Pol II) at nucleotide resolution have provided key insights into the underlying molecular mechanisms that regulate transcriptional responses to stress. In addition, recent kinetic analyses of transcriptional control under heat stress have shown how cells 'prewire' and rapidly execute genome-wide changes in transcription while concurrently becoming poised for recovery. The regulation of Pol II at genes and enhancers in response to heat stress is coupled to chromatin modification and compartmentalization, as well as to co-transcriptional RNA processing. These mechanistic features seem to apply broadly to other coordinated genome-regulatory responses.
Conflict of interest statement
The authors declare no competing interests.
Figures


References
-
- Richter K, Haslbeck M, Buchner J. The heat shock response: life on the verge of death. Mol Cell. 2010;40:253–266. - PubMed
-
- Walter P, Ron D. The unfolded protein response: from stress pathway to homeostatic regulation. Science. 2011;334:1081–1086. - PubMed
-
- Quirós PM, Langer T, López-Otín C. New roles for mitochondrial proteases in health, ageing and disease. Nat Rev Mol Cell Biol. 2015;16:345–359. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
