

The Long Pentraxin 3 Plays a Role in Bone Turnover and Repair
- PMID: 29556234
- PMCID: PMC5845433
- DOI: 10.3389/fimmu.2018.00417
The Long Pentraxin 3 Plays a Role in Bone Turnover and Repair
Abstract
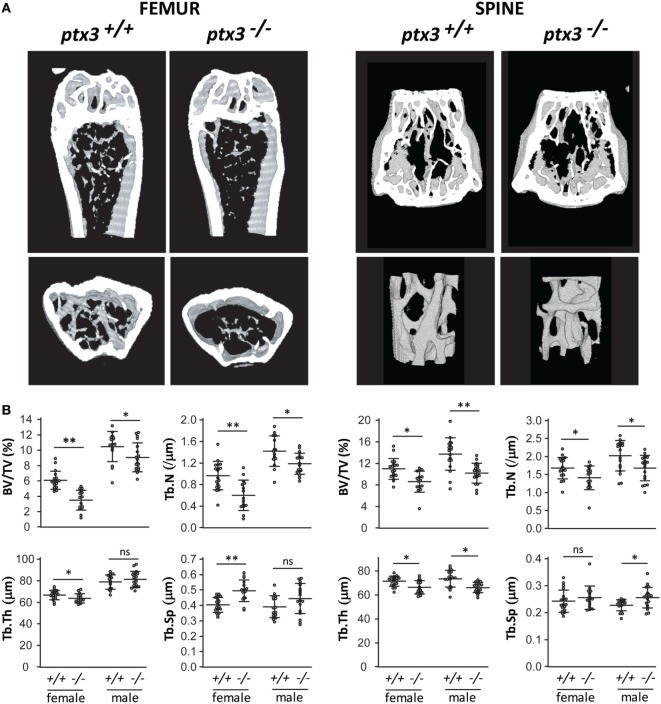
Pentraxin 3 (PTX3) is an inflammatory mediator acting as a fluid-phase pattern recognition molecule and playing an essential role in innate immunity and matrix remodeling. Inflammatory mediators also contribute to skeletal homeostasis, operating at multiple levels in physiological and pathological conditions. This study was designed to investigate the role of PTX3 in physiological skeletal remodeling and bone healing. Micro-computed tomography (μCT) and bone histomorphometry of distal femur showed that PTX3 gene-targeted female and male mice (ptx3-/- ) had lower trabecular bone volume than their wild-type (ptx3+/+ ) littermates (BV/TV by μCT: 3.50 ± 1.31 vs 6.09 ± 1.17 for females, p < 0.0001; BV/TV 9.06 ± 1.89 vs 10.47 ± 1.97 for males, p = 0.0435). In addition, μCT revealed lower trabecular bone volume in second lumbar vertebra of ptx3-/- mice. PTX3 was increasingly expressed during osteoblast maturation in vitro and was able to reverse the negative effect of fibroblast growth factor 2 (FGF2) on osteoblast differentiation. This effect was specific for the N-terminal domain of PTX3 that contains the FGF2-binding site. By using the closed transversal tibial fracture model, we found that ptx3-/- female mice formed significantly less mineralized callus during the anabolic phase following fracture injury compared to ptx3+/+ mice (BV/TV 17.05 ± 4.59 vs 20.47 ± 3.32, p = 0.0195). Non-hematopoietic periosteal cells highly upregulated PTX3 expression during the initial phase of fracture healing, particularly CD51+ and αSma+ osteoprogenitor subsets, and callus tissue exhibited concomitant expression of PTX3 and FGF2 around the fracture site. Thus, PTX3 supports maintenance of the bone mass possibly by inhibiting FGF2 and its negative impact on bone formation. Moreover, PTX3 enables timely occurring sequence of callus mineralization after bone fracture injury. These results indicate that PTX3 plays an important role in bone homeostasis and in proper matrix mineralization during fracture repair, a reflection of the function of this molecule in tissue homeostasis and repair.
Keywords: bone healing; bone turnover; fibroblast growth factor 2; inflammation; osteoblast; osteoclast; pentraxin 3.
Figures







References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous