

The Subthalamic Nucleus: Unravelling New Roles and Mechanisms in the Control of Action
- PMID: 29557710
- PMCID: PMC7263363
- DOI: 10.1177/1073858418763594
The Subthalamic Nucleus: Unravelling New Roles and Mechanisms in the Control of Action
Abstract
How do we decide what we do? This is the essence of action control, the process of selecting the most appropriate response among multiple possible choices. Suboptimal action control can involve a failure to initiate or adapt actions, or conversely it can involve making actions impulsively. There has been an increasing focus on the specific role of the subthalamic nucleus (STN) in action control. This has been fueled by the clinical relevance of this basal ganglia nucleus as a target for deep brain stimulation (DBS), primarily in Parkinson's disease but also in obsessive-compulsive disorder. The context of DBS has opened windows to study STN function in ways that link neuroscientific and clinical fields closely together, contributing to an exceptionally high level of two-way translation. In this review, we first outline the role of the STN in both motor and nonmotor action control, and then discuss how these functions might be implemented by neuronal activity in the STN. Gaining a better understanding of these topics will not only provide important insights into the neurophysiology of action control but also the pathophysiological mechanisms relevant for several brain disorders and their therapies.
Keywords: action control; decision making; deep brain stimulation; neuronal oscillations; subthalamic nucleus.
Conflict of interest statement
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Figures
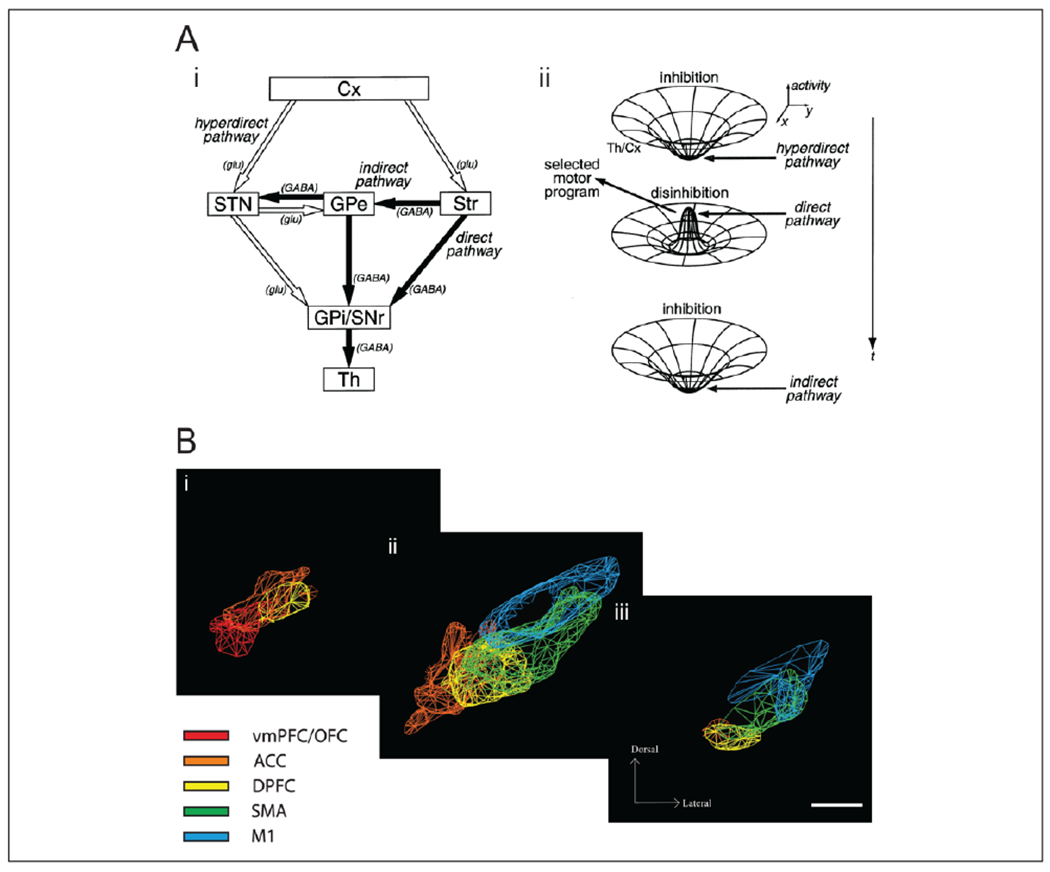
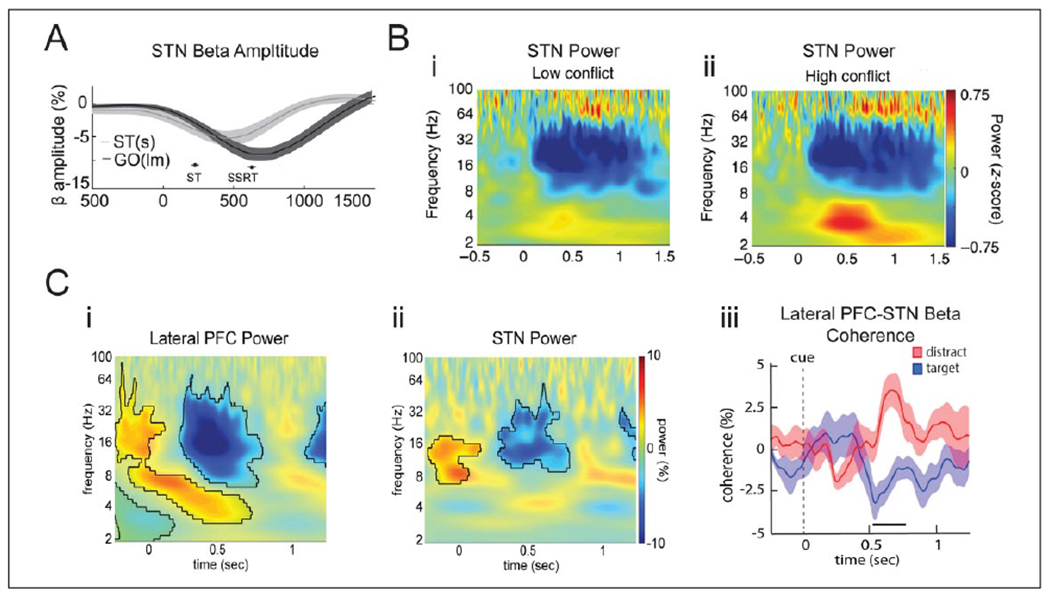
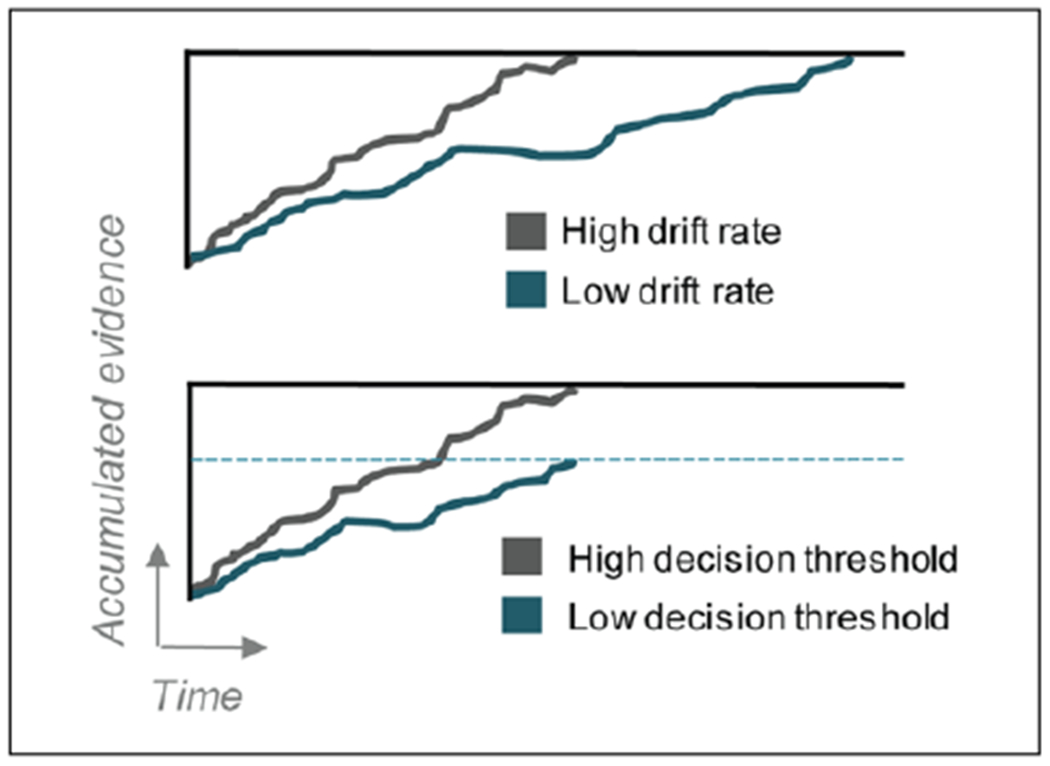
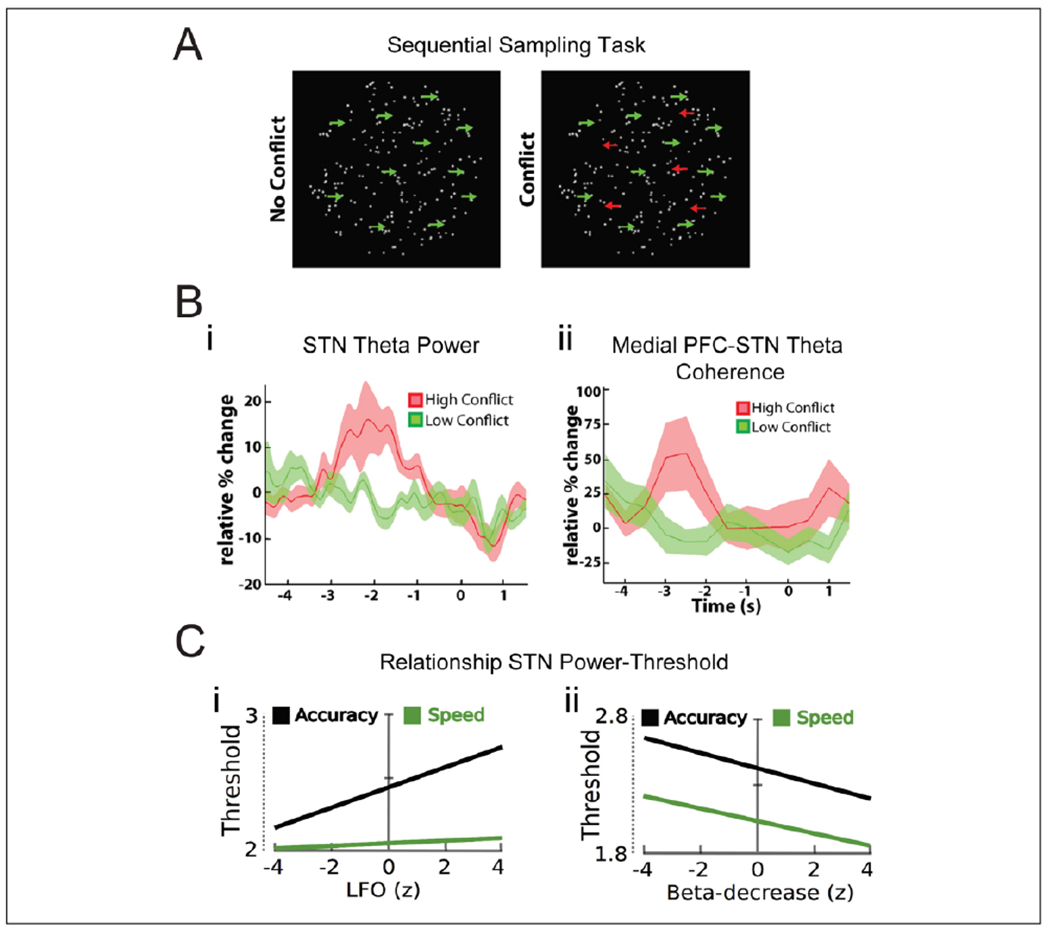
References
-
- Alegre M, Lopez-Azcarate J, Obeso I, Wilkinson L, Rodriguez-Oroz MC, Valencia M, and others 2013. The subthalamic nucleus is involved in successful inhibition in the stop-signal task: a local field potential study in Parkinson’s disease. Exp Neurol 239:1–12. - PubMed
-
- Alexander GE, Crutcher MD, DeLong MR. 1990. Basal ganglia-thalamocortical circuits: parallel substrates for motor, oculomotor, “prefrontal” and “limbic” functions. Prog Brain Res 85:119–46. - PubMed
-
- Alexander GE, DeLong MR, Strick PL. 1986. Parallel organization of functionally segregated circuits linking basal ganglia and cortex. Annu Rev Neurosci 9:357–81. - PubMed
-
- Arcos D, Sierra A, Nunez A, Flores G, Aceves J, Arias-Montano JA. 2003. Noradrenaline increases the firing rate of a subpopulation of rat subthalamic neurones through the activation of alpha 1-adrenoceptors. Neuropharmacology 45(8):1070–9. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
