

Bordetella pertussis pertactin knock-out strains reveal immunomodulatory properties of this virulence factor
- PMID: 29559630
- PMCID: PMC5861065
- DOI: 10.1038/s41426-018-0039-8
Bordetella pertussis pertactin knock-out strains reveal immunomodulatory properties of this virulence factor
Abstract
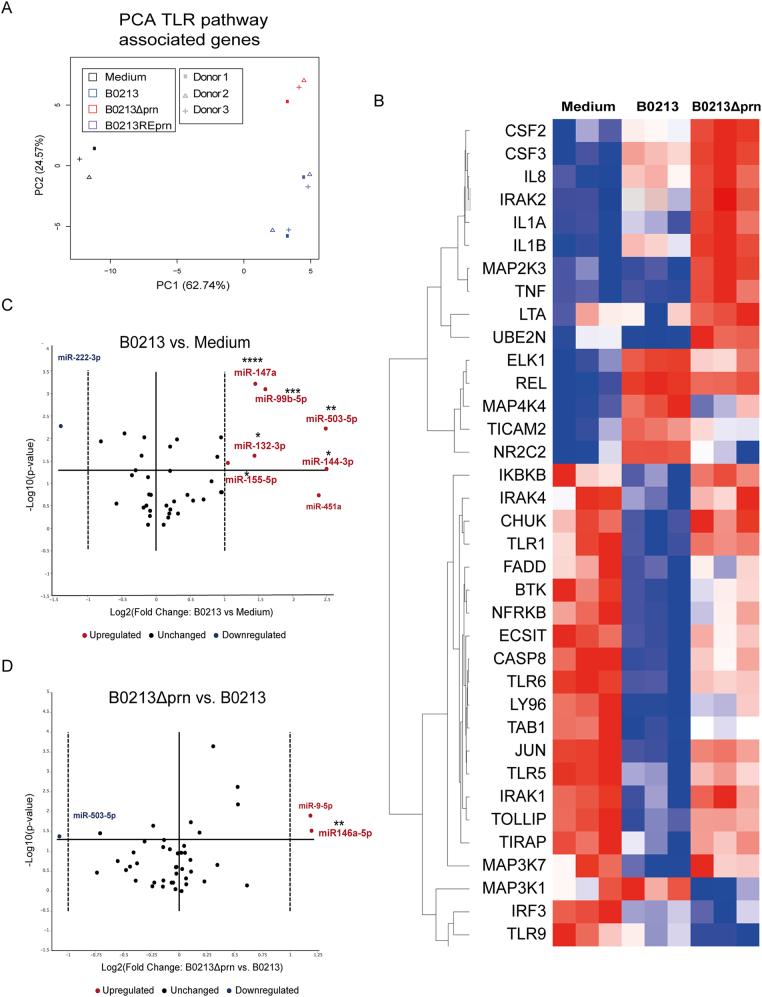
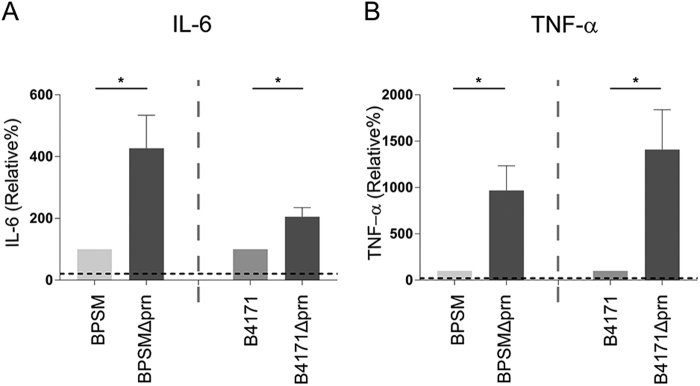
Whooping cough, caused by Bordetella pertussis, has resurged and presents a global health burden worldwide. B. pertussis strains unable to produce the acellular pertussis vaccine component pertactin (Prn), have been emerging and in some countries represent up to 95% of recent clinical isolates. Knowledge on the effect that Prn deficiency has on infection and immunity to B. pertussis is crucial for the development of new strategies to control this disease. Here, we characterized the effect of Prn production by B. pertussis on human and murine dendritic cell (DC) maturation as well as in a murine model for pertussis infection. We incubated human monocyte-derived DCs (moDCs) with multiple isogenic Prn knockout (Prn-KO) and corresponding parental B. pertussis strains constructed either in laboratory reference strains with a Tohama I background or in a recently circulating clinical isolate. Results indicate that, compared to the parental strains, Prn-KO strains induced an increased production of pro-inflammatory cytokines by moDCs. This pro-inflammatory phenotype was also observed upon stimulation of murine bone marrow-derived DCs. Moreover, RNA sequencing analysis of lungs from mice infected with B. pertussis Prn-KO revealed increased expression of genes involved in cell death. These in vitro and in vivo findings indicate that B. pertussis strains which do not produce Prn induce a stronger pro-inflammatory response and increased cell death upon infection, suggesting immunomodulatory properties for Prn.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
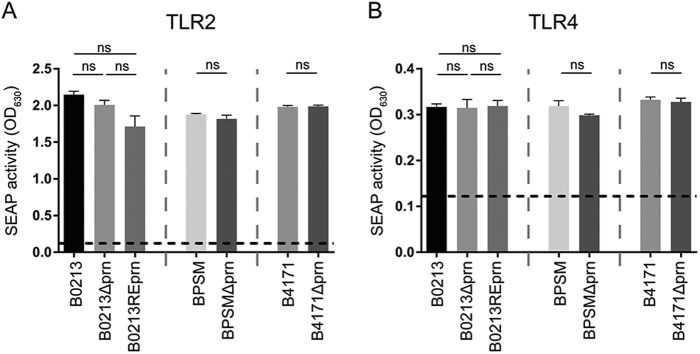
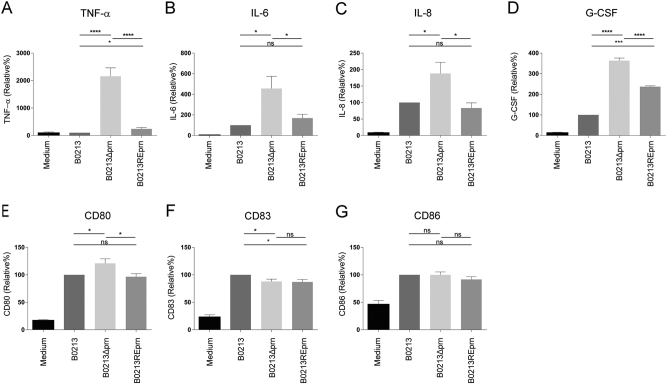
Similar articles
-
Pertactin negative Bordetella pertussis demonstrates higher fitness under vaccine selection pressure in a mixed infection model.Vaccine. 2015 Nov 17;33(46):6277-81. doi: 10.1016/j.vaccine.2015.09.064. Epub 2015 Oct 2. Vaccine. 2015. PMID: 26432908
-
Pertactin-negative Bordetella pertussis strains: evidence for a possible selective advantage.Clin Infect Dis. 2015 Jan 15;60(2):223-7. doi: 10.1093/cid/ciu788. Epub 2014 Oct 9. Clin Infect Dis. 2015. PMID: 25301209
-
Pertactin-Negative and Filamentous Hemagglutinin-Negative Bordetella pertussis, Australia, 2013-2017.Emerg Infect Dis. 2019 Jun;25(6):1196-1199. doi: 10.3201/eid2506.180240. Emerg Infect Dis. 2019. PMID: 31107218 Free PMC article.
-
Pertactin deficiency of Bordetella pertussis: Insights into epidemiology, and perspectives on surveillance and public health impact.Hum Vaccin Immunother. 2024 Dec 31;20(1):2435134. doi: 10.1080/21645515.2024.2435134. Epub 2024 Dec 17. Hum Vaccin Immunother. 2024. PMID: 39686838 Free PMC article.
-
Bordetella pertussis and pertactin-deficient clinical isolates: lessons for pertussis vaccines.Expert Rev Vaccines. 2014 Sep;13(9):1135-46. doi: 10.1586/14760584.2014.932254. Epub 2014 Jun 23. Expert Rev Vaccines. 2014. PMID: 24953157 Review.
Cited by
-
Genomic dissection of the microevolution of Australian epidemic Bordetella pertussis.Emerg Microbes Infect. 2022 Dec;11(1):1460-1473. doi: 10.1080/22221751.2022.2077129. Emerg Microbes Infect. 2022. PMID: 35543519 Free PMC article.
-
Modulation of Inflammatory Signaling Molecules in Bordetella pertussis Antigen-Challenged Human Monocytes in Presence of Adrenergic Agonists.Vaccines (Basel). 2022 Feb 17;10(2):321. doi: 10.3390/vaccines10020321. Vaccines (Basel). 2022. PMID: 35214778 Free PMC article.
-
Generating enhanced mucosal immunity against Bordetella pertussis: current challenges and new directions.Front Immunol. 2023 Feb 21;14:1126107. doi: 10.3389/fimmu.2023.1126107. eCollection 2023. Front Immunol. 2023. PMID: 36895562 Free PMC article.
-
Live attenuated pertussis vaccine for prevention and treatment of allergic airway inflammation in mice.NPJ Vaccines. 2022 Jun 23;7(1):66. doi: 10.1038/s41541-022-00494-w. NPJ Vaccines. 2022. PMID: 35739108 Free PMC article.
-
Non-specific Effects of Live Attenuated Pertussis Vaccine Against Heterologous Infectious and Inflammatory Diseases.Front Immunol. 2018 Dec 7;9:2872. doi: 10.3389/fimmu.2018.02872. eCollection 2018. Front Immunol. 2018. PMID: 30581436 Free PMC article. Review.
References
-
- Olin P, Rasmussen F, Gustafsson L, Hallander HO, Heijbel H. Randomised controlled trial of two-component, three-component, and five-component acellular pertussis vaccines compared with whole-cell pertussis vaccine. Ad Hoc Group for the Study of Pertussis Vaccines. Lancet. 1997;350:1569–1577. doi: 10.1016/S0140-6736(97)06508-2. - DOI - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials