

De novo variants in CAMK2A and CAMK2B cause neurodevelopmental disorders
- PMID: 29560374
- PMCID: PMC5846454
- DOI: 10.1002/acn3.528
De novo variants in CAMK2A and CAMK2B cause neurodevelopmental disorders
Abstract
Objective: α (CAMK2A) and β (CAMK2B) isoforms of Calcium/calmodulin-dependent protein kinase II (CaMKII) play a pivotal role in neuronal plasticity and in learning and memory processes in the brain. Here, we explore the possible involvement of α- and β-CaMKII variants in neurodevelopmental disorders.
Methods: Whole-exome sequencing was performed for 976 individuals with intellectual disability, developmental delay, and epilepsy. The effect of CAMK2A and CAMK2B variants on CaMKII structure and firing of neurons was evaluated by computational structural analysis, immunoblotting, and electrophysiological analysis.
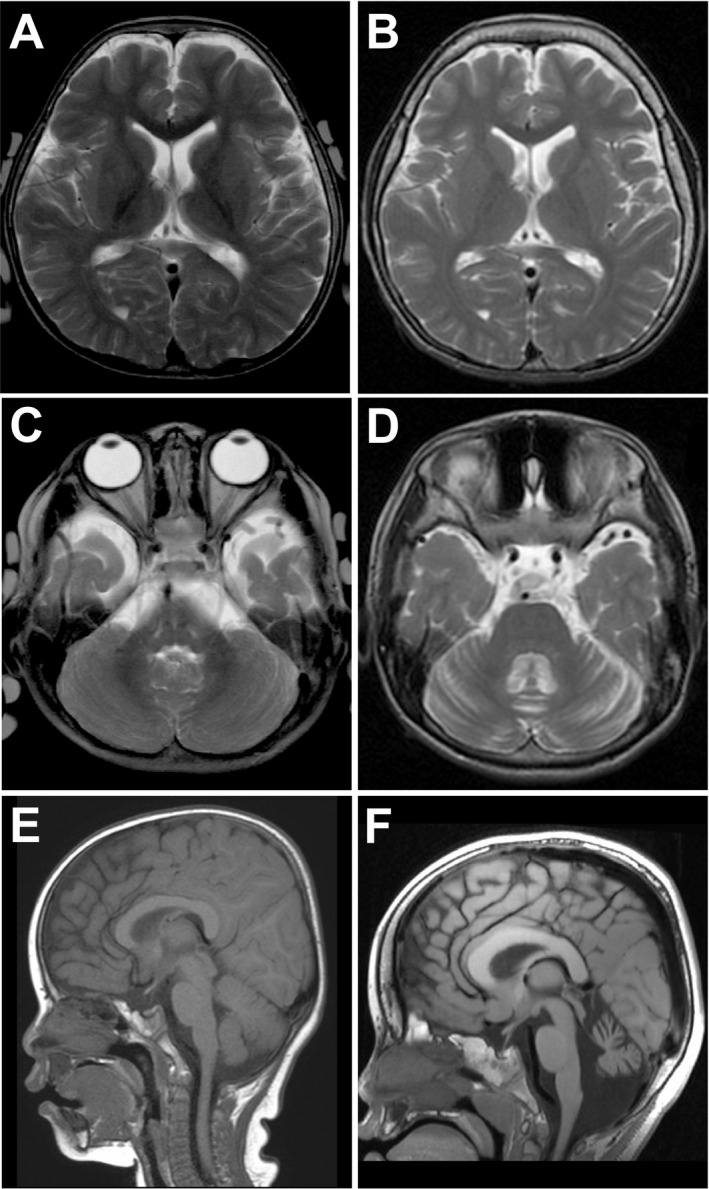
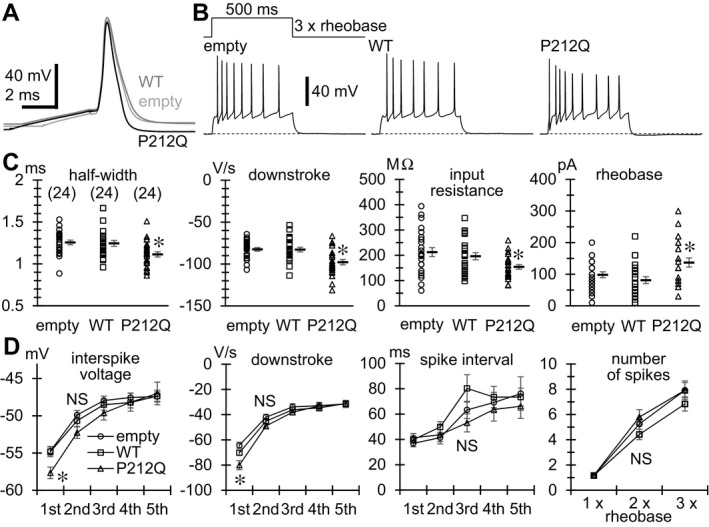
Results: We identified a total of five de novo CAMK2A and CAMK2B variants in three and two individuals, respectively. Seizures were common to three individuals with CAMK2A variants. Using a minigene splicing assay, we demonstrated that a splice site variant caused skipping of exon 11 leading to an in-frame deletion of the regulatory segment of CaMKII α. By structural analysis, four missense variants are predicted to impair the interaction between the kinase domain and the regulatory segment responsible for the autoinhibition of its kinase activity. The Thr286/Thr287 phosphorylation as a result of release from autoinhibition was increased in three mutants when the mutants were stably expressed in Neuro-2a neuroblastoma cells. Expression of a CaMKII α mutant in primary hippocampal neurons significantly increased A-type K+ currents, which facilitated spike repolarization of single action potentials.
Interpretation: Our data highlight the importance of CaMKII α and CaMKII β and their autoinhibitory regulation in human brain function, and suggest the enhancement of A-type K+ currents as a possible pathophysiological basis.
Figures





References
-
- Liu XB, Murray KD. Neuronal excitability and calcium/calmodulin‐dependent protein kinase type II: location, location, location. Epilepsia 2012;53(Suppl 1):45–52. - PubMed
-
- Yang E, Schulman H. Structural examination of autoregulation of multifunctional calcium/calmodulin‐dependent protein kinase II. J Biol Chem 1999;274:26199–26208. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
