

Targeting potassium channels to treat cerebellar ataxia
- PMID: 29560375
- PMCID: PMC5846455
- DOI: 10.1002/acn3.527
Targeting potassium channels to treat cerebellar ataxia
Abstract
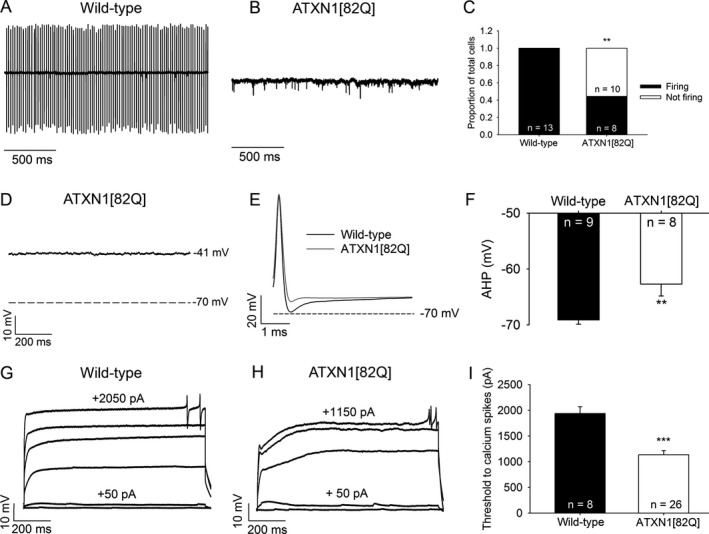
Objective: Purkinje neuron dysfunction is associated with cerebellar ataxia. In a mouse model of spinocerebellar ataxia type 1 (SCA1), reduced potassium channel function contributes to altered membrane excitability resulting in impaired Purkinje neuron spiking. We sought to determine the relationship between altered membrane excitability and motor dysfunction in SCA1 mice.
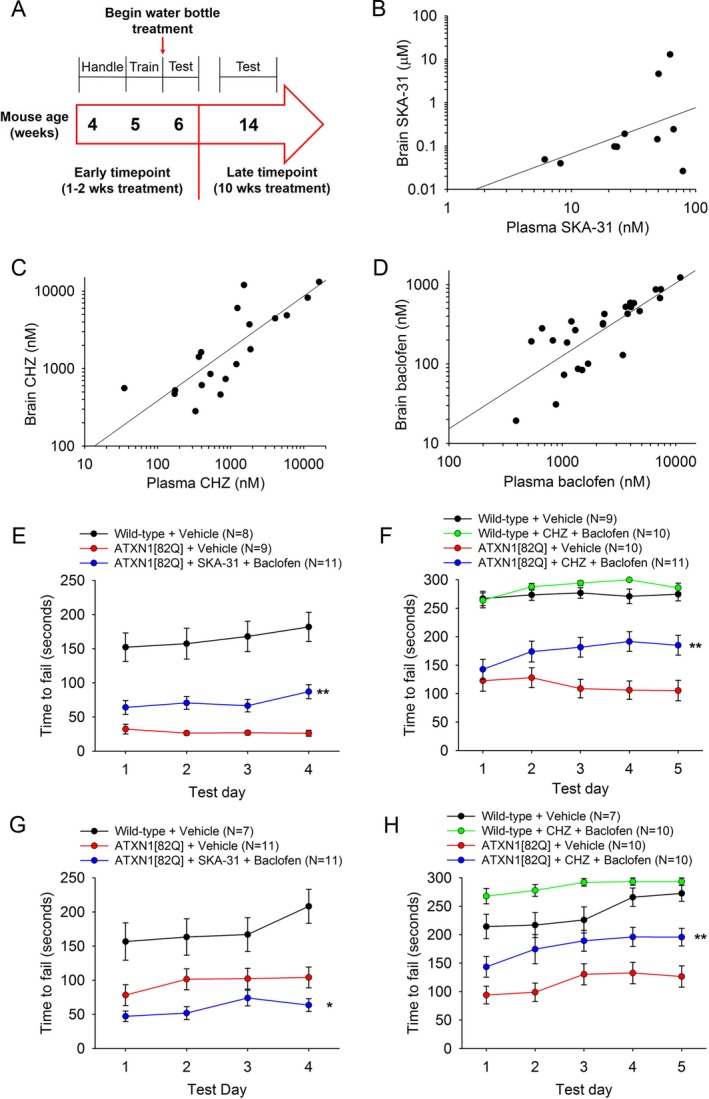
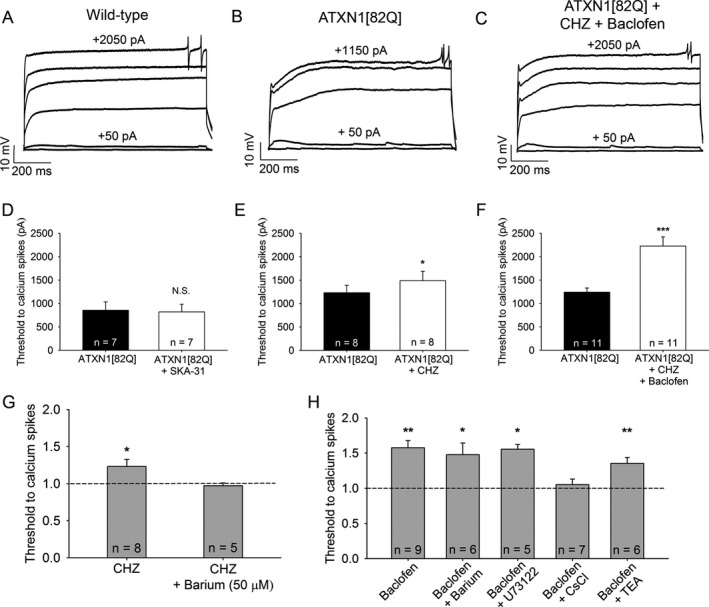
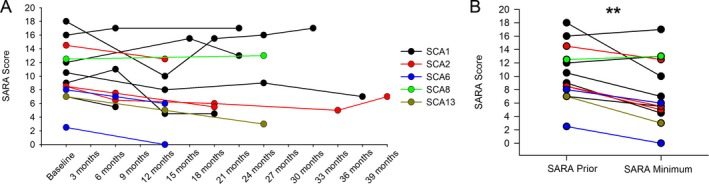
Methods: Patch-clamp recordings in acute cerebellar slices and motor phenotype testing were used to identify pharmacologic agents which improve Purkinje neuron physiology and motor performance in SCA1 mice. Additionally, we retrospectively reviewed records of patients with SCA1 and other autosomal-dominant SCAs with prominent Purkinje neuron involvement to determine whether currently approved potassium channel activators were tolerated.
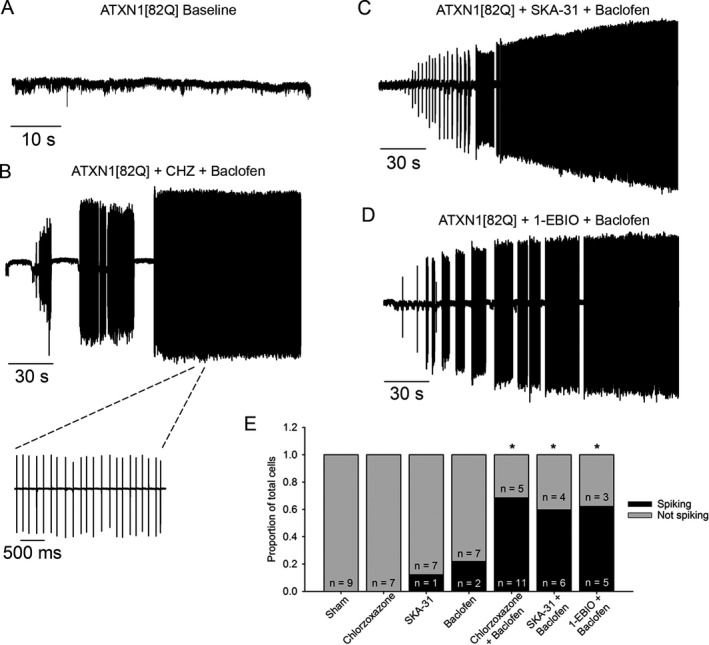
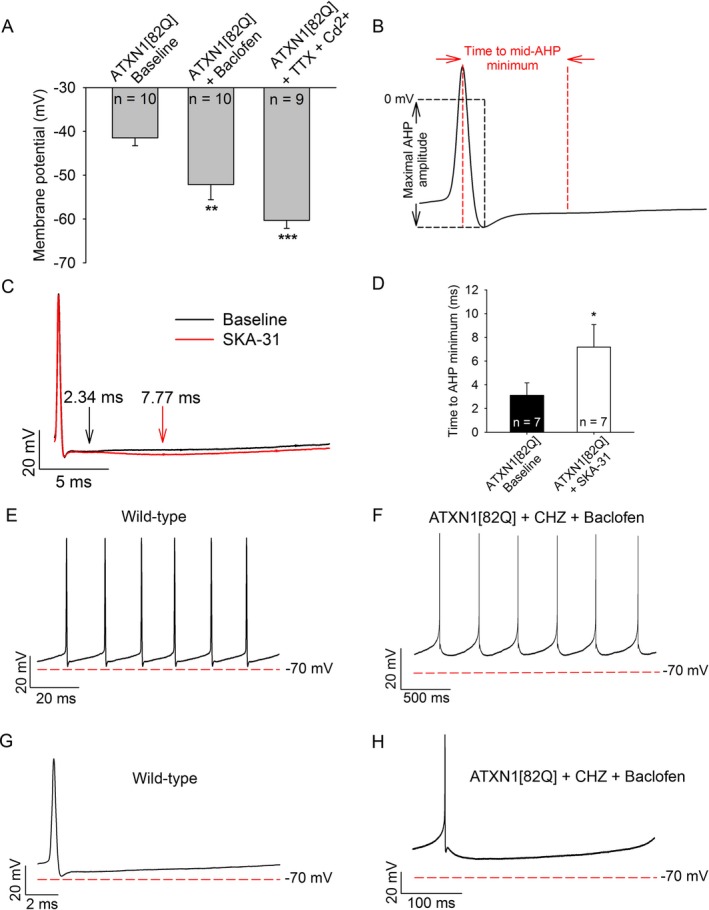
Results: Activating calcium-activated and subthreshold-activated potassium channels improved Purkinje neuron spiking impairment in SCA1 mice (P < 0.05). Additionally, dendritic hyperexcitability was improved by activating subthreshold-activated potassium channels but not calcium-activated potassium channels (P < 0.01). Improving spiking and dendritic hyperexcitability through a combination of chlorzoxazone and baclofen produced sustained improvements in motor dysfunction in SCA1 mice (P < 0.01). Retrospective review of SCA patient records suggests that co-treatment with chlorzoxazone and baclofen is tolerated.
Interpretation: Targeting both altered spiking and dendritic membrane excitability is associated with sustained improvements in motor performance in SCA1 mice, while targeting altered spiking alone produces only short-term improvements in motor dysfunction. Potassium channel activators currently in clinical use are well tolerated and may provide benefit in SCA patients. Future clinical trials with potassium channel activators are warranted in cerebellar ataxia.
Figures
References
-
- Ferrer I, Genis D, Davalos A, et al. The Purkinje cell in olivopontocerebellar atrophy. A Golgi and immunocytochemical study. Neuropathol Appl Neurobiol 1994;20:38–46. - PubMed
