

Cdk1 phosphorylation of Esp1/Separase functions with PP2A and Slk19 to regulate pericentric Cohesin and anaphase onset
- PMID: 29561844
- PMCID: PMC5880407
- DOI: 10.1371/journal.pgen.1007029
Cdk1 phosphorylation of Esp1/Separase functions with PP2A and Slk19 to regulate pericentric Cohesin and anaphase onset
Abstract
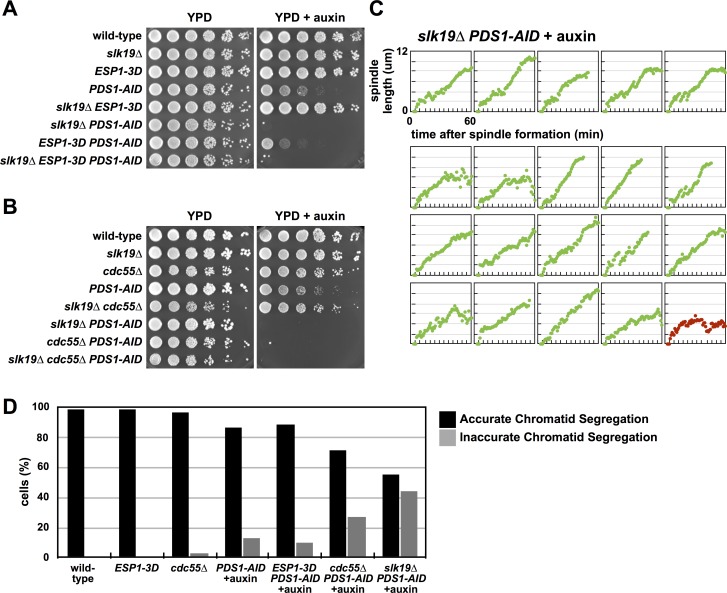
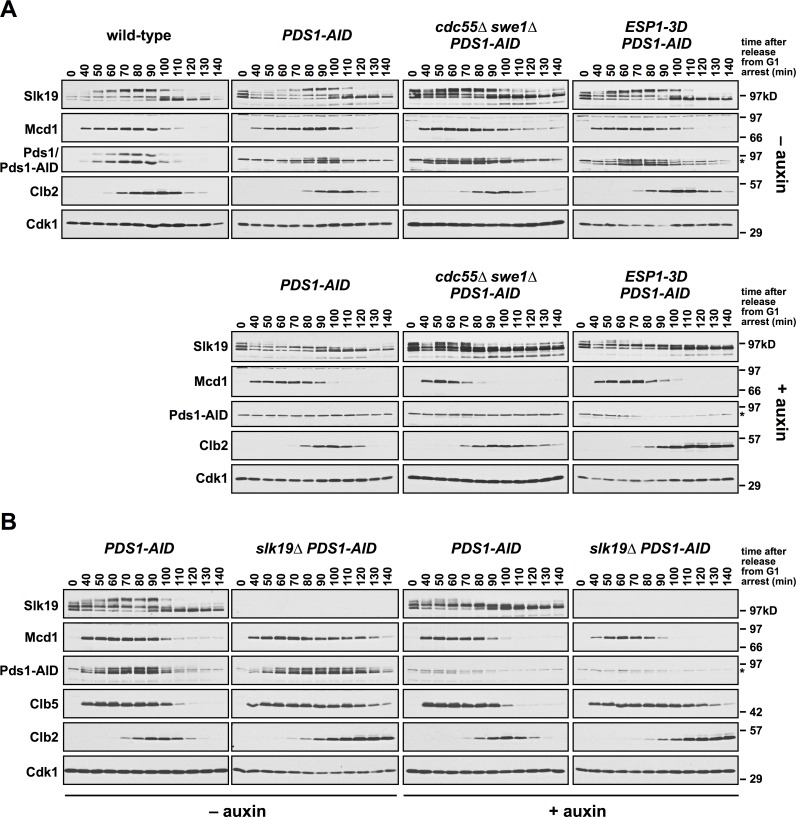
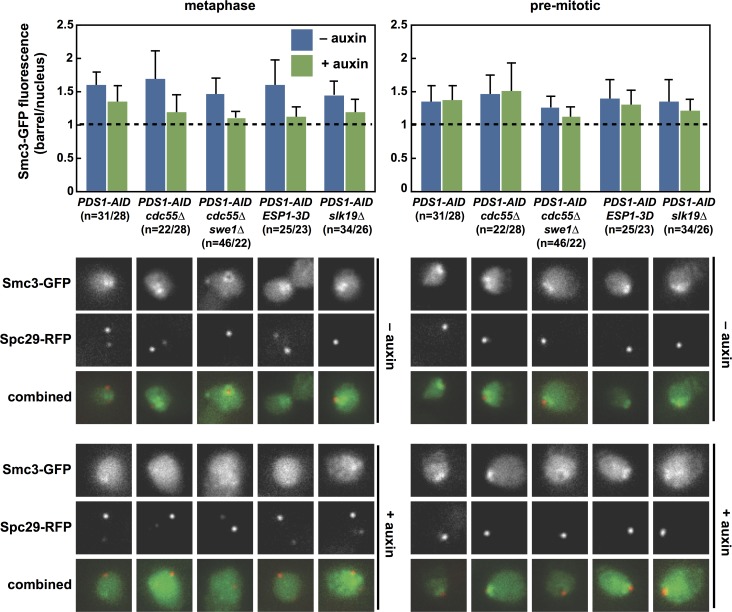
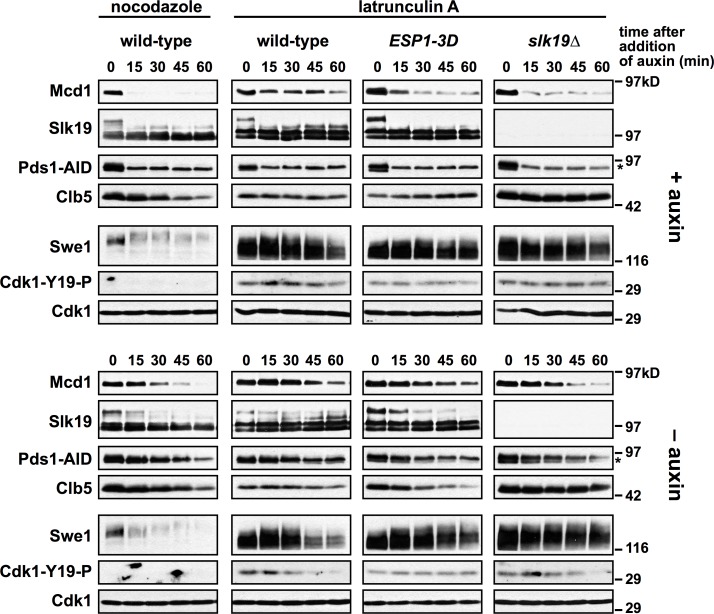
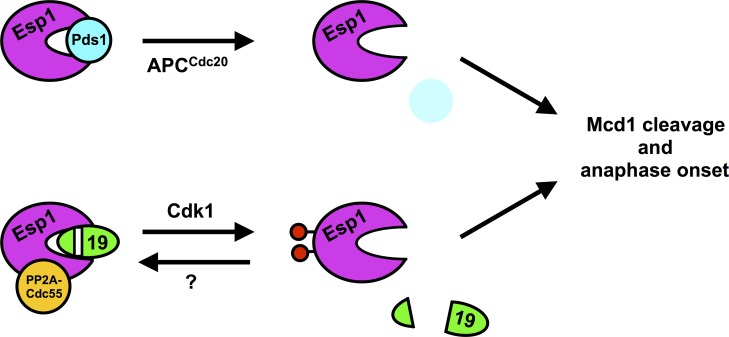
Anaphase onset is an irreversible cell cycle transition that is triggered by the activation of the protease Separase. Separase cleaves the Mcd1 (also known as Scc1) subunit of Cohesin, a complex of proteins that physically links sister chromatids, triggering sister chromatid separation. Separase is regulated by the degradation of the anaphase inhibitor Securin which liberates Separase from inhibitory Securin/Separase complexes. In many organisms, Securin is not essential suggesting that Separase is regulated by additional mechanisms. In this work, we show that in budding yeast Cdk1 activates Separase (Esp1 in yeast) through phosphorylation to trigger anaphase onset. Esp1 activation is opposed by protein phosphatase 2A associated with its regulatory subunit Cdc55 (PP2ACdc55) and the spindle protein Slk19. Premature anaphase spindle elongation occurs when Securin (Pds1 in yeast) is inducibly degraded in cells that also contain phospho-mimetic mutations in ESP1, or deletion of CDC55 or SLK19. This striking phenotype is accompanied by advanced degradation of Mcd1, disruption of pericentric Cohesin organization and chromosome mis-segregation. Our findings suggest that PP2ACdc55 and Slk19 function redundantly with Pds1 to inhibit Esp1 within pericentric chromatin, and both Pds1 degradation and Cdk1-dependent phosphorylation of Esp1 act together to trigger anaphase onset.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

Similar articles
-
PP2ACdc55 dephosphorylates Pds1 and inhibits spindle elongation in S. cerevisiae.J Cell Sci. 2020 Jul 29;133(14):jcs243766. doi: 10.1242/jcs.243766. J Cell Sci. 2020. PMID: 32591482 Free PMC article.
-
Pds1/Esp1-dependent and -independent sister chromatid separation in mutants defective for protein phosphatase 2A.Proc Natl Acad Sci U S A. 2006 Oct 31;103(44):16290-5. doi: 10.1073/pnas.0607856103. Epub 2006 Oct 18. Proc Natl Acad Sci U S A. 2006. PMID: 17050679 Free PMC article.
-
A role for the budding yeast separase, Esp1, in Ty1 element retrotransposition.PLoS Genet. 2015 Mar 30;11(3):e1005109. doi: 10.1371/journal.pgen.1005109. eCollection 2015 Mar. PLoS Genet. 2015. PMID: 25822502 Free PMC article.
-
Cell cycle mechanisms of sister chromatid separation; roles of Cut1/separin and Cut2/securin.Genes Cells. 2000 Jan;5(1):1-8. doi: 10.1046/j.1365-2443.2000.00306.x. Genes Cells. 2000. PMID: 10651900 Review.
-
Separase and Roads to Disengage Sister Chromatids during Anaphase.Int J Mol Sci. 2023 Feb 27;24(5):4604. doi: 10.3390/ijms24054604. Int J Mol Sci. 2023. PMID: 36902034 Free PMC article. Review.
Cited by
-
Kif17 phosphorylation regulates photoreceptor outer segment turnover.BMC Cell Biol. 2018 Nov 20;19(1):25. doi: 10.1186/s12860-018-0177-9. BMC Cell Biol. 2018. PMID: 30458707 Free PMC article.
-
PP2ACdc55 dephosphorylates Pds1 and inhibits spindle elongation in S. cerevisiae.J Cell Sci. 2020 Jul 29;133(14):jcs243766. doi: 10.1242/jcs.243766. J Cell Sci. 2020. PMID: 32591482 Free PMC article.
-
Mapping separase-mediated cleavage in situ.NAR Genom Bioinform. 2022 Nov 18;4(4):lqac085. doi: 10.1093/nargab/lqac085. eCollection 2022 Dec. NAR Genom Bioinform. 2022. PMID: 36415827 Free PMC article.
-
Slk19 enhances cross-linking of microtubules by Ase1 and Stu1.Mol Biol Cell. 2021 Nov 1;32(21):ar22. doi: 10.1091/mbc.E21-05-0279. Epub 2021 Sep 8. Mol Biol Cell. 2021. PMID: 34495712 Free PMC article.
-
Characterization of a novel separase-interacting protein and candidate new securin, Eip1p, in the fungal pathogen Candida albicans.Mol Biol Cell. 2019 Sep 1;30(19):2469-2489. doi: 10.1091/mbc.E18-11-0696. Epub 2019 Aug 14. Mol Biol Cell. 2019. PMID: 31411946 Free PMC article.
References
-
- Michaelis C, Ciosk R, Nasmyth K. Cohesins: chromosomal proteins that prevent premature separation of sister chromatids. Cell. 1997;91: 35–45. - PubMed
-
- Gruber S, Haering CH, Nasmyth K. Chromosomal cohesin forms a ring. Cell.2003;112: 765–777. - PubMed
-
- Haering CH, Farcas A-M, Arumugam P, Metson J, Nasmyth K. The cohesin ring concatenates sister DNA molecules. Nature. 2008;454: 297–301. doi: 10.1038/nature07098 - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
