

Insulin resistance in cavefish as an adaptation to a nutrient-limited environment
- PMID: 29562229
- PMCID: PMC5989729
- DOI: 10.1038/nature26136
Insulin resistance in cavefish as an adaptation to a nutrient-limited environment
Erratum in
-
Author Correction: Insulin resistance in cavefish as an adaptation to a nutrient-limited environment.Nature. 2020 Dec;588(7836):E1. doi: 10.1038/s41586-020-2953-x. Nature. 2020. PMID: 33188369
Abstract
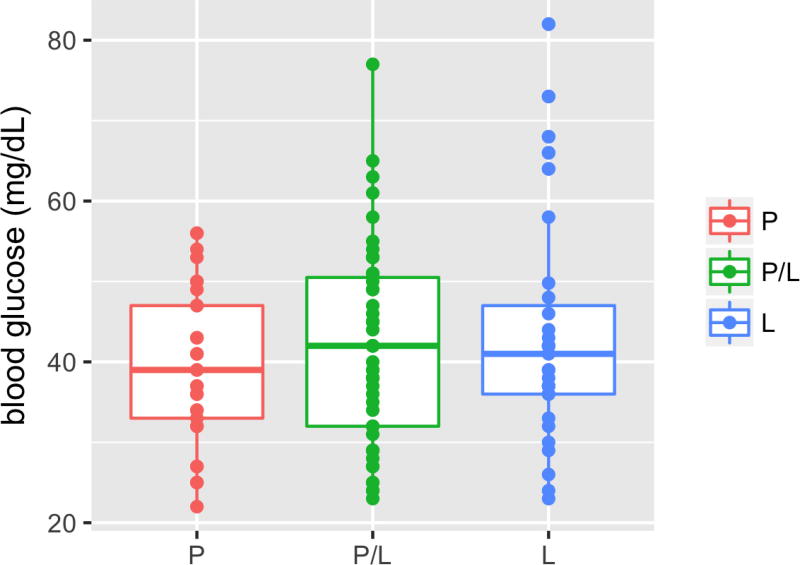
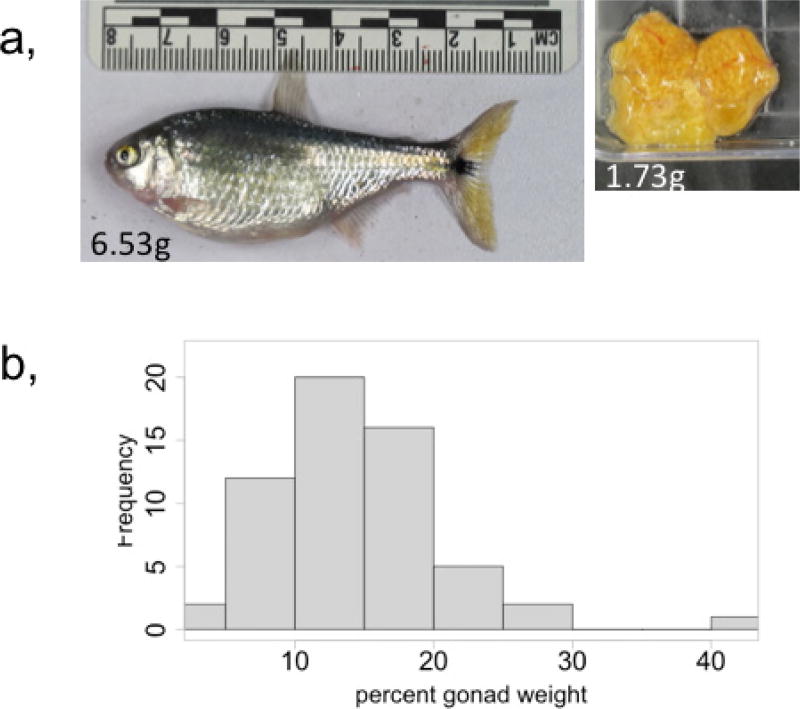
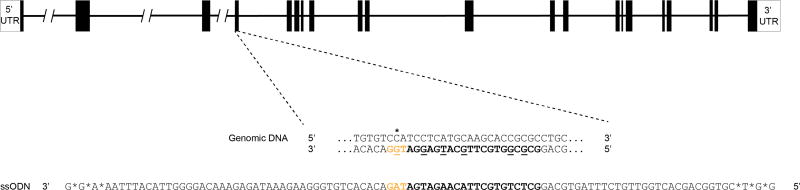
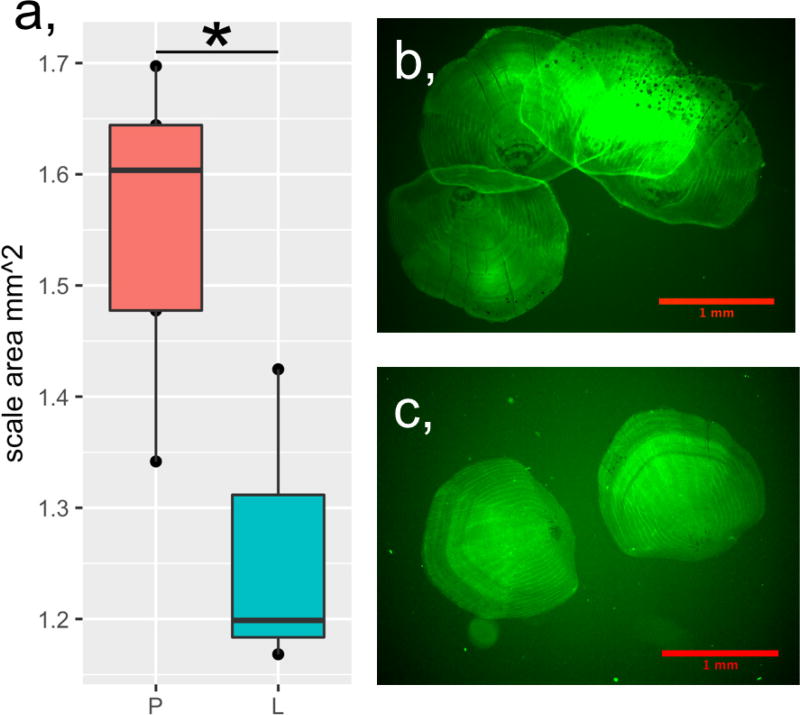
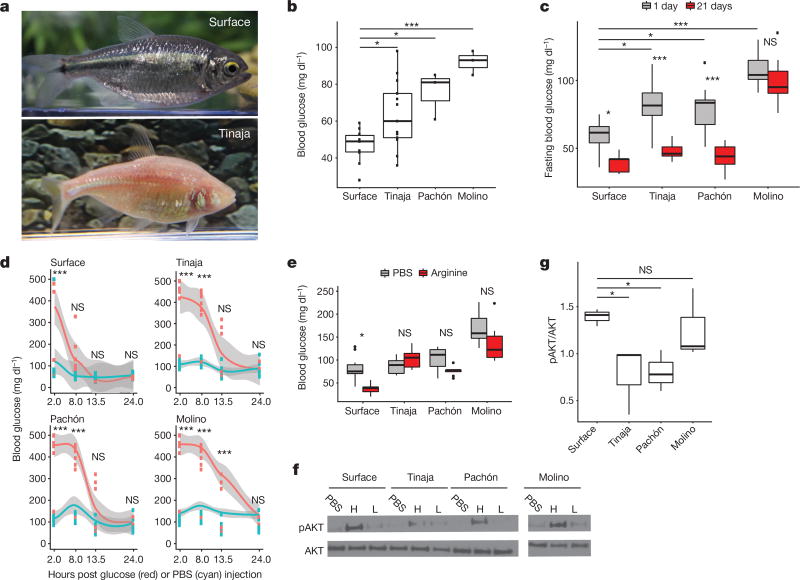
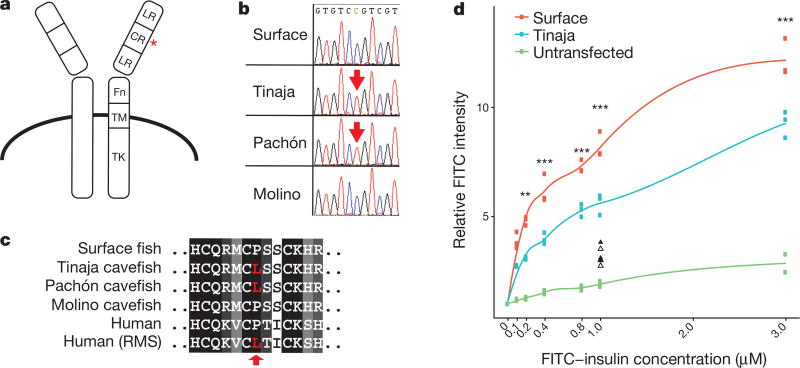
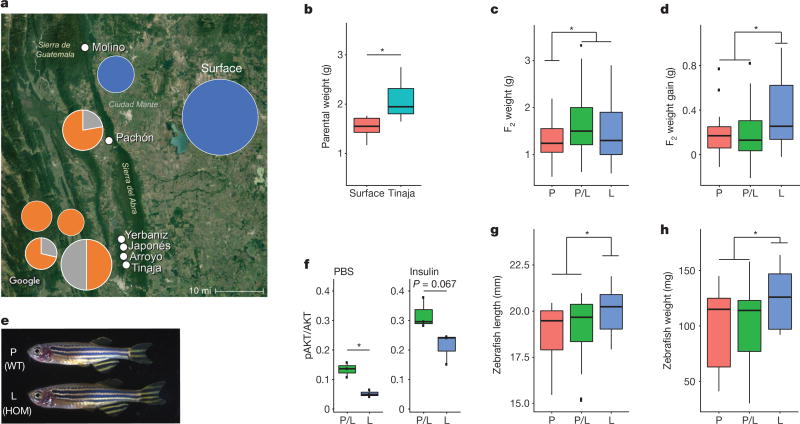
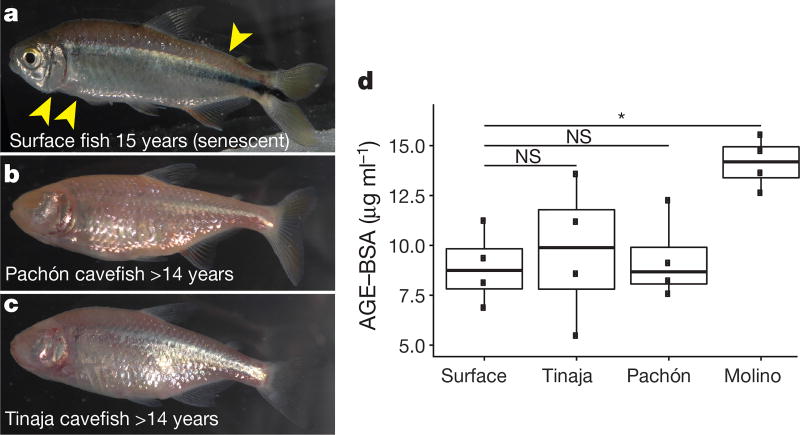
Periodic food shortages are a major challenge faced by organisms in natural habitats. Cave-dwelling animals must withstand long periods of nutrient deprivation, as-in the absence of photosynthesis-caves depend on external energy sources such as seasonal floods. Here we show that cave-adapted populations of the Mexican tetra, Astyanax mexicanus, have dysregulated blood glucose homeostasis and are insulin-resistant compared to river-adapted populations. We found that multiple cave populations carry a mutation in the insulin receptor that leads to decreased insulin binding in vitro and contributes to hyperglycaemia. Hybrid fish from surface-cave crosses carrying this mutation weigh more than non-carriers, and zebrafish genetically engineered to carry the mutation have increased body weight and insulin resistance. Higher body weight may be advantageous in caves as a strategy to cope with an infrequent food supply. In humans, the identical mutation in the insulin receptor leads to a severe form of insulin resistance and reduced lifespan. However, cavefish have a similar lifespan to surface fish and do not accumulate the advanced glycation end-products in the blood that are typically associated with the progression of diabetes-associated pathologies. Our findings suggest that diminished insulin signalling is beneficial in a nutrient-limited environment and that cavefish may have acquired compensatory mechanisms that enable them to circumvent the typical negative effects associated with failure to regulate blood glucose levels.
Conflict of interest statement
The authors declare no competing interests.
Figures
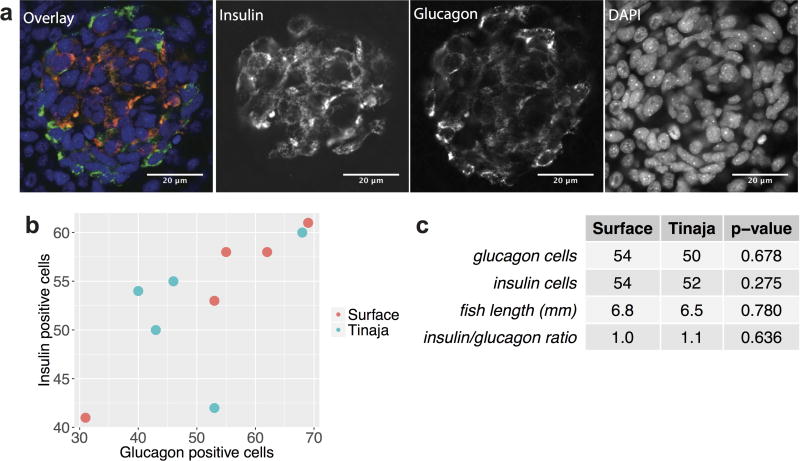
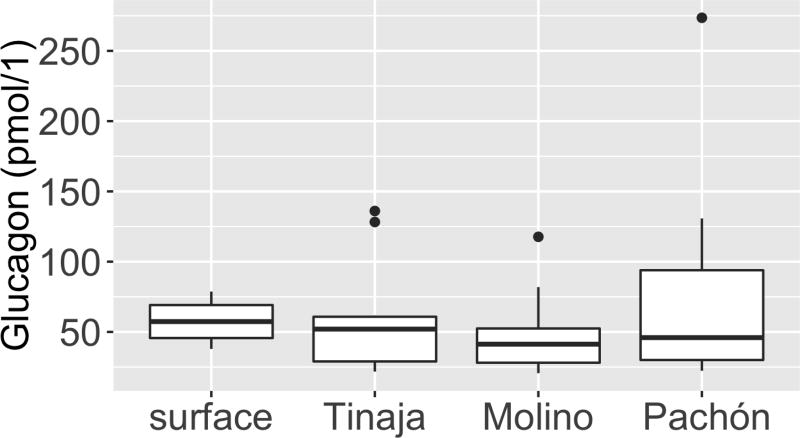
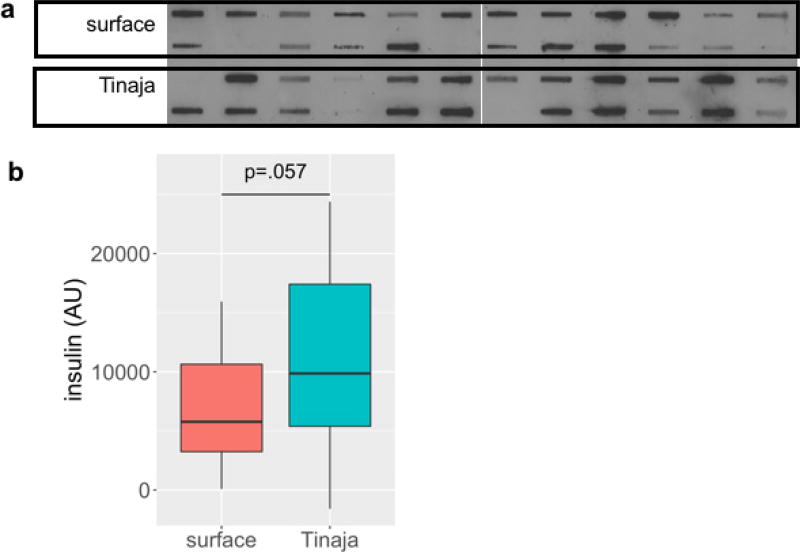
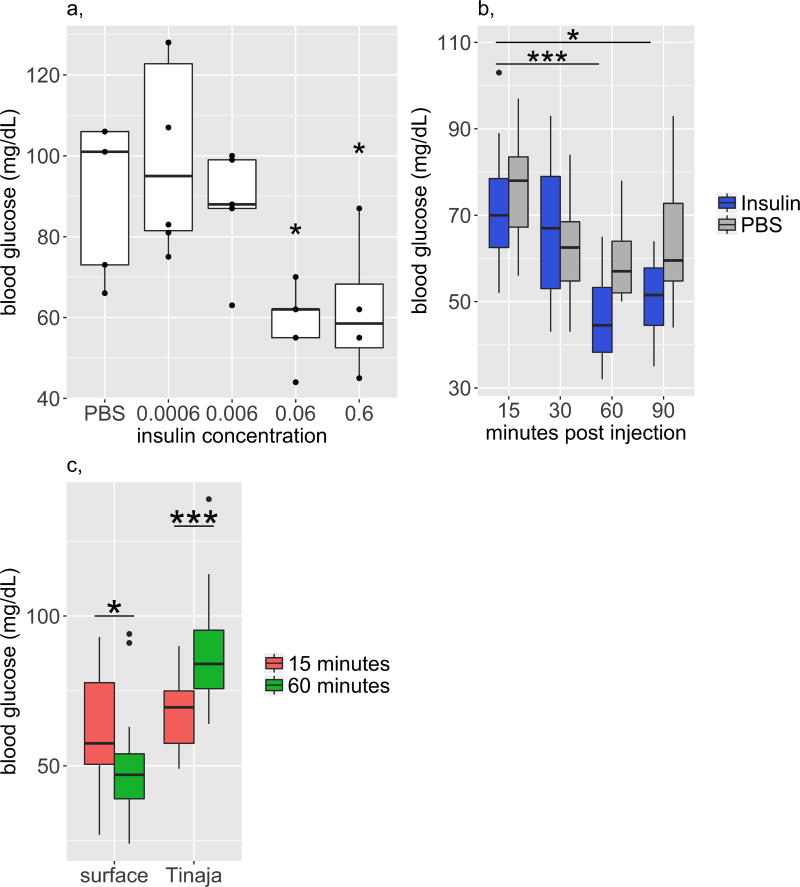
Comment in
-
The healthy diabetic cavefish conundrum.Nature. 2018 Mar 29;555(7698):595-597. doi: 10.1038/d41586-018-03242-0. Nature. 2018. PMID: 29595777 No abstract available.
References
-
- Culver DC, Pipan T. The Biology of Caves and Other Subterranean Habitats. Oxford Univ. Press; 2009.
-
- Horst Wilkens US. Evolution in the Dark, Darwin’s Loss Without Selection. Springer; 2017.
-
- Hüppop K. Oxygen consumption of Astyanax fasciatus (Characidae, Pisces): a comparison of epigean and hypogean populations. Environ. Biol. Fishes. 1986;17:299–308.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
