

Structural insights into the voltage and phospholipid activation of the mammalian TPC1 channel
- PMID: 29562233
- PMCID: PMC5886804
- DOI: 10.1038/nature26139
Structural insights into the voltage and phospholipid activation of the mammalian TPC1 channel
Abstract
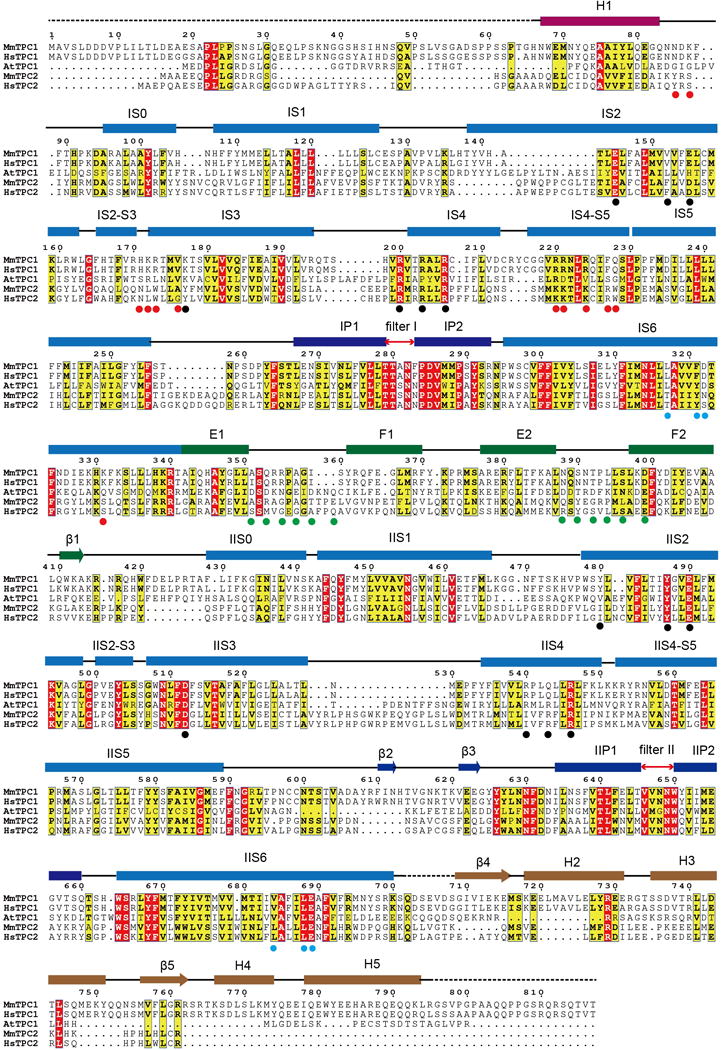
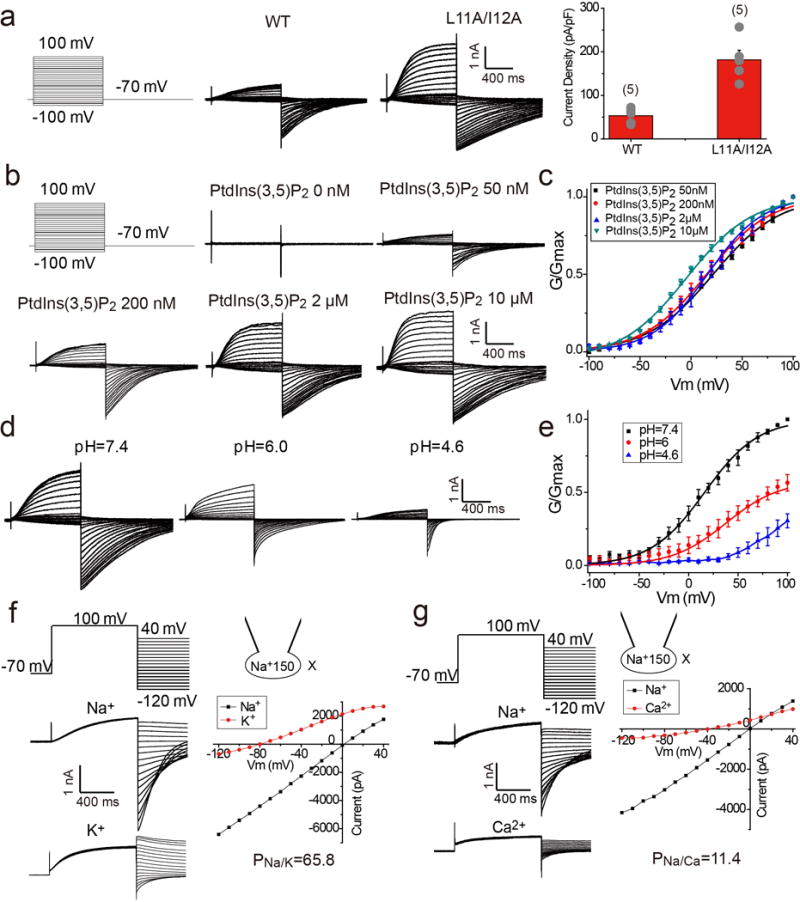
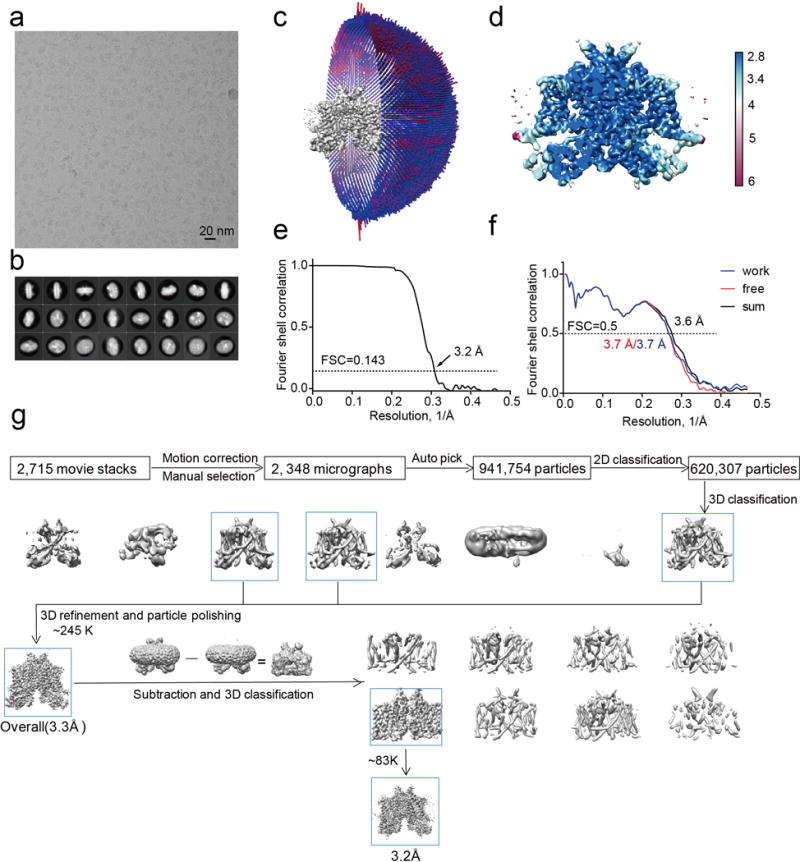
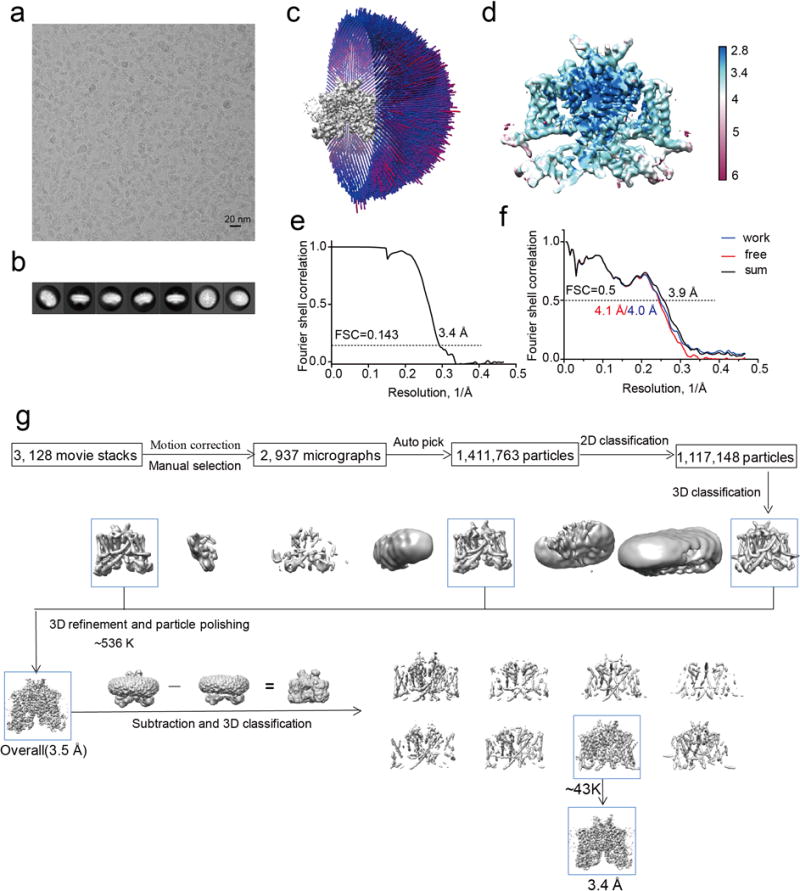
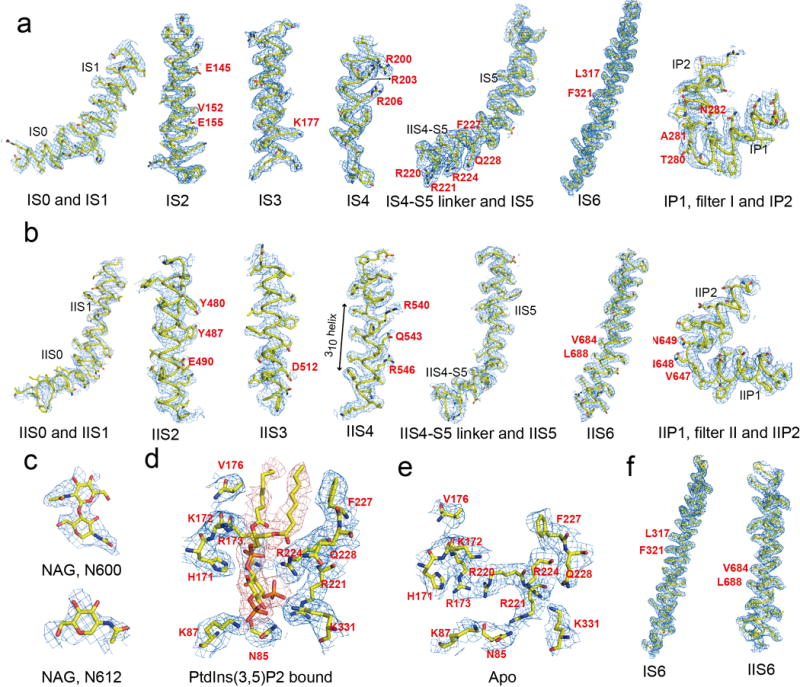
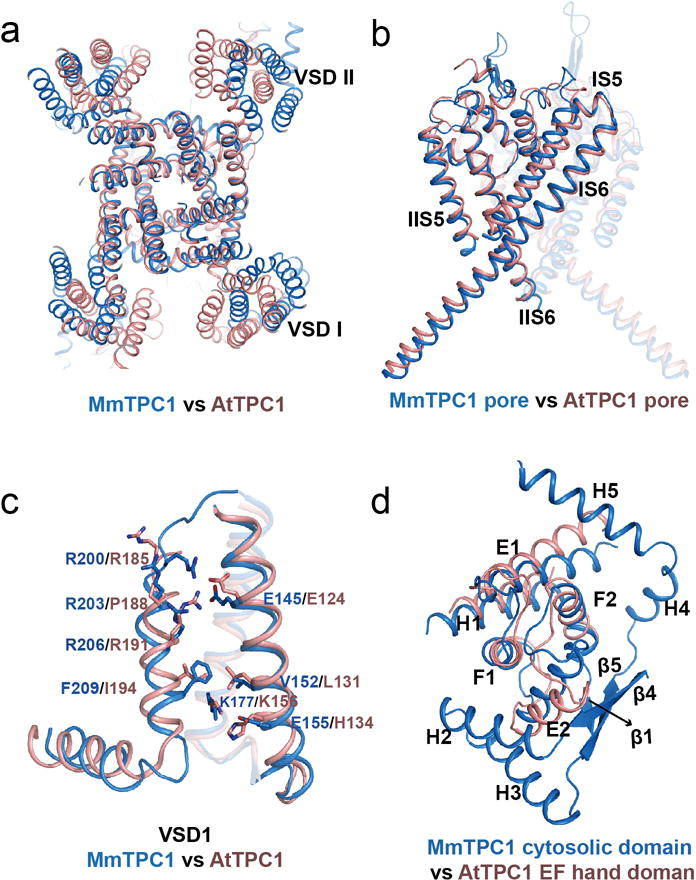
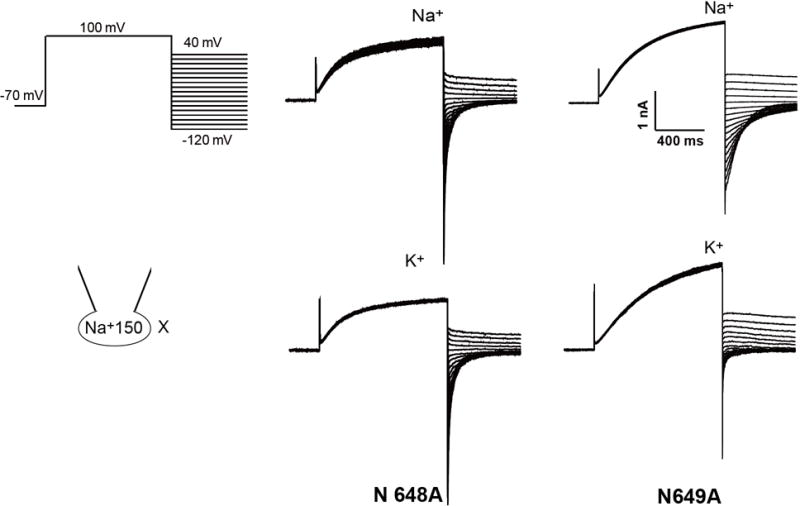
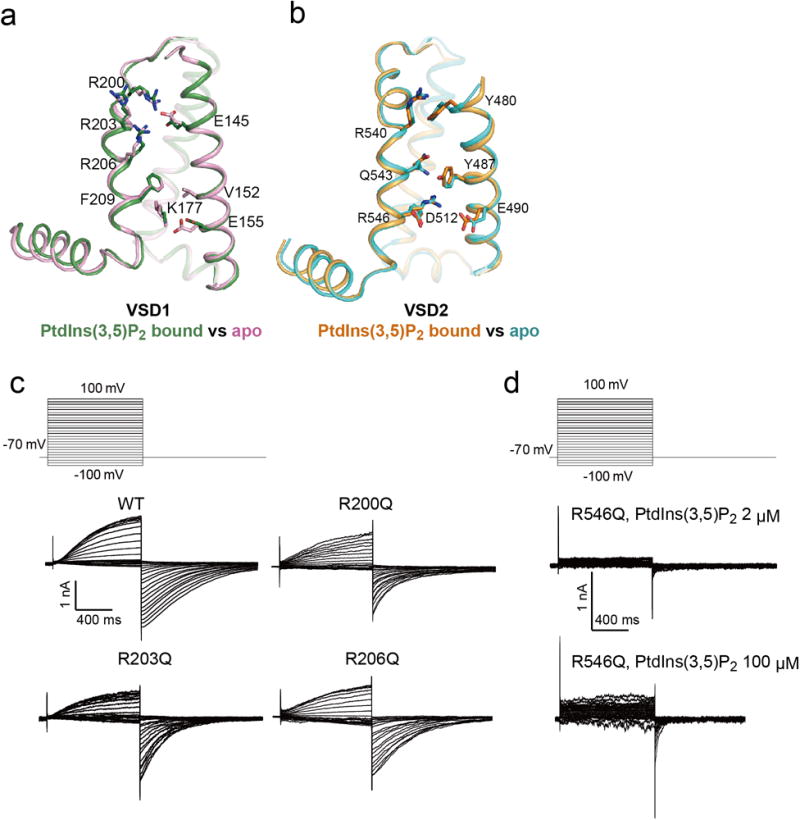
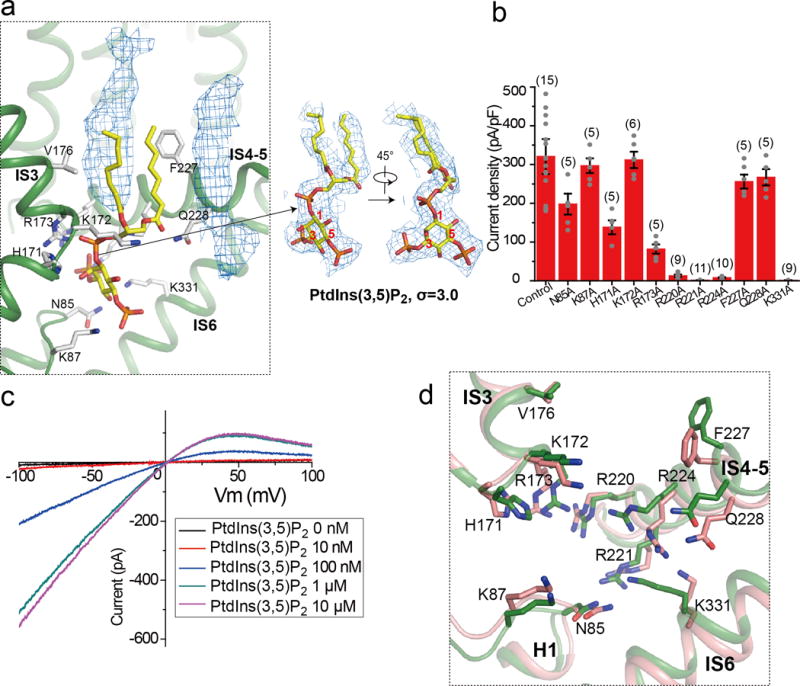
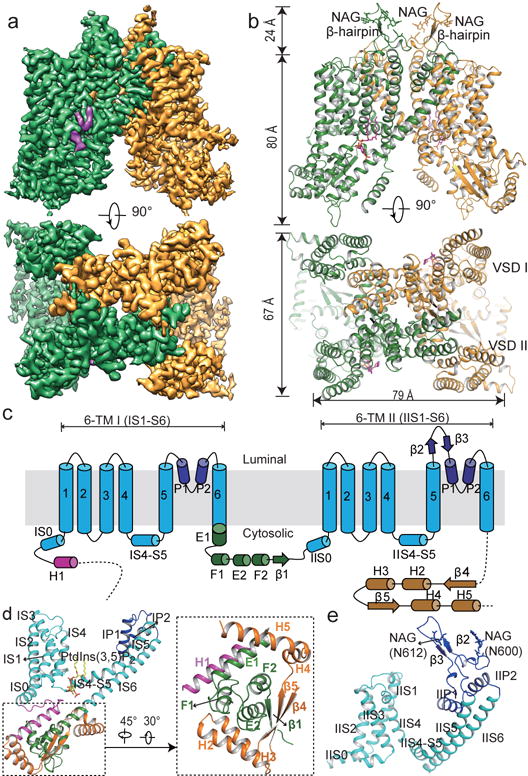
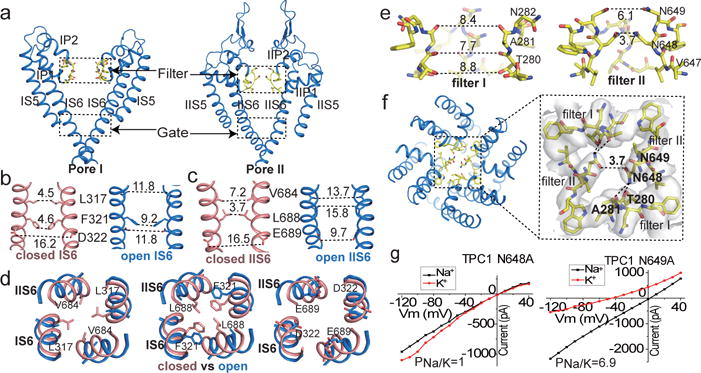
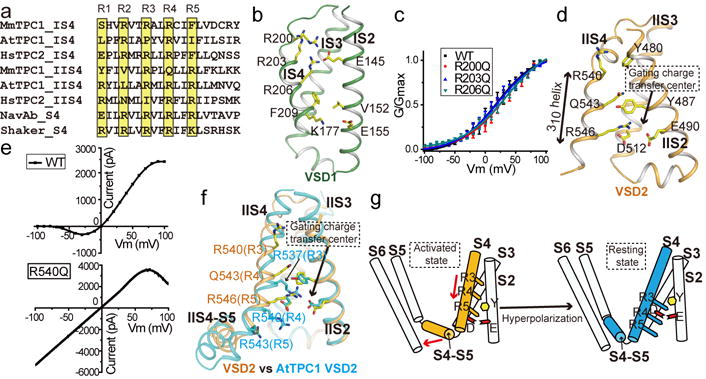
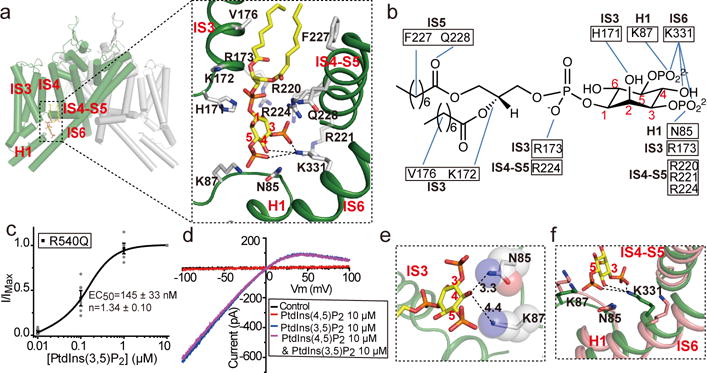
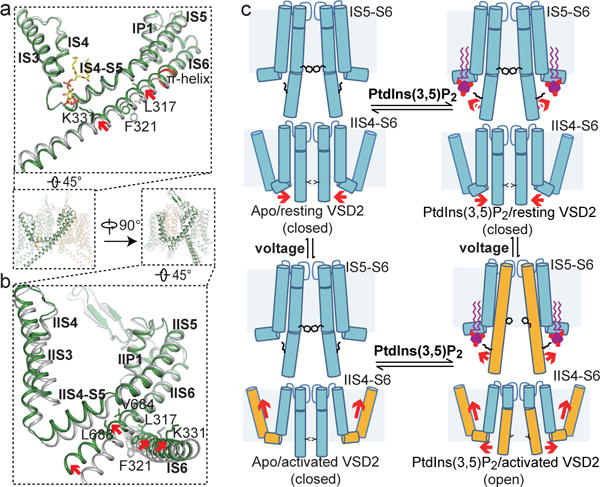
The organellar two-pore channel (TPC) functions as a homodimer, in which each subunit contains two homologous Shaker-like six-transmembrane (6-TM)-domain repeats. TPCs belong to the voltage-gated ion channel superfamily and are ubiquitously expressed in animals and plants. Mammalian TPC1 and TPC2 are localized at the endolysosomal membrane, and have critical roles in regulating the physiological functions of these acidic organelles. Here we present electron cryo-microscopy structures of mouse TPC1 (MmTPC1)-a voltage-dependent, phosphatidylinositol 3,5-bisphosphate (PtdIns(3,5)P2)-activated Na+-selective channel-in both the apo closed state and ligand-bound open state. Combined with functional analysis, these structures provide comprehensive structural insights into the selectivity and gating mechanisms of mammalian TPC channels. The channel has a coin-slot-shaped ion pathway in the filter that defines the selectivity of mammalian TPCs. Only the voltage-sensing domain from the second 6-TM domain confers voltage dependence on MmTPC1. Endolysosome-specific PtdIns(3,5)P2 binds to the first 6-TM domain and activates the channel under conditions of depolarizing membrane potential. Structural comparisons between the apo and PtdIns(3,5)P2-bound structures show the interplay between voltage and ligand in channel activation. These MmTPC1 structures reveal lipid binding and regulation in a 6-TM voltage-gated channel, which is of interest in light of the emerging recognition of the importance of phosphoinositide regulation of ion channels.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
Comment in
-
Two-pore channels open up.Nature. 2018 Apr 5;556(7699):38-40. doi: 10.1038/d41586-018-02783-8. Nature. 2018. PMID: 29620760 No abstract available.
References
-
- Furuichi T, Cunningham KW, Muto S. A putative two pore channel AtTPC1 mediates Ca(2+) flux in Arabidopsis leaf cells. Plant Cell Physiol. 2001;42:900–905. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
