

Novel Aspects of Polynucleotide Phosphorylase Function in Streptomyces
- PMID: 29562650
- PMCID: PMC5872136
- DOI: 10.3390/antibiotics7010025
Novel Aspects of Polynucleotide Phosphorylase Function in Streptomyces
Abstract
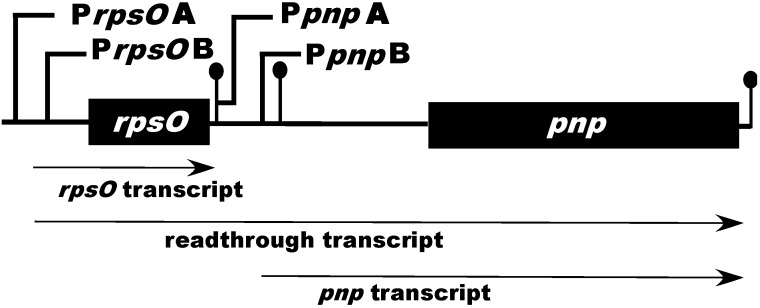
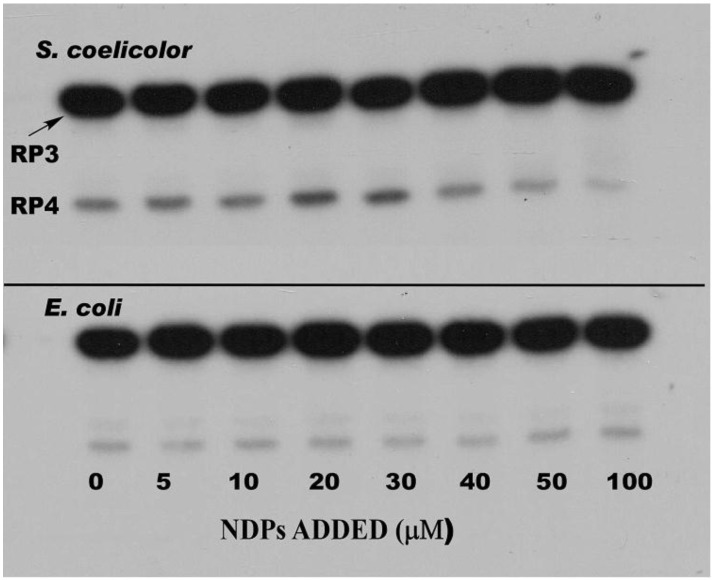
Polynucleotide phosphorylase (PNPase) is a 3'-5'-exoribnuclease that is found in most bacteria and in some eukaryotic organelles. The enzyme plays a key role in RNA decay in these systems. PNPase structure and function have been studied extensively in Escherichiacoli, but there are several important aspects of PNPase function in Streptomyces that differ from what is observed in E. coli and other bacterial genera. This review highlights several of those differences: (1) the organization and expression of the PNPase gene in Streptomyces; (2) the possible function of PNPase as an RNA 3'-polyribonucleotide polymerase in Streptomyces; (3) the function of PNPase as both an exoribonuclease and as an RNA 3'-polyribonucleotide polymerase in Streptomyces; (4) the function of (p)ppGpp as a PNPase effector in Streptomyces. The review concludes with a consideration of a number of unanswered questions regarding the function of Streptomyces PNPase, which can be examined experimentally.
Keywords: (p)ppGpp; RNA decay; Streptomyces; antibiotic; polyadenylation; polynucleotide phosphorylase; promoter; regulation; ribonuclease.
Conflict of interest statement
The author declares no conflicts of interest.
Figures




References
-
- Grunberg-Manago M., Ochoa S. Enzymatic synthesis and breakdown of polynucleotides: Polynucleotide phosphorylase. J. Am. Chem. Soc. 1955;77:3165–3166. doi: 10.1021/ja01616a093. - DOI
-
- Godefroy-Colburn T., Grunberg-Manago M. Polynucleotide phosphorylase. Enzymes. 1972;7:533–574.
-
- Littauer U.Z., Soreq H. Polynucleotide phosphorylase. Enzymes. 1982;15:517–553.
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
