

Microbiome-host systems interactions: protective effects of propionate upon the blood-brain barrier
- PMID: 29562936
- PMCID: PMC5863458
- DOI: 10.1186/s40168-018-0439-y
Microbiome-host systems interactions: protective effects of propionate upon the blood-brain barrier
Abstract
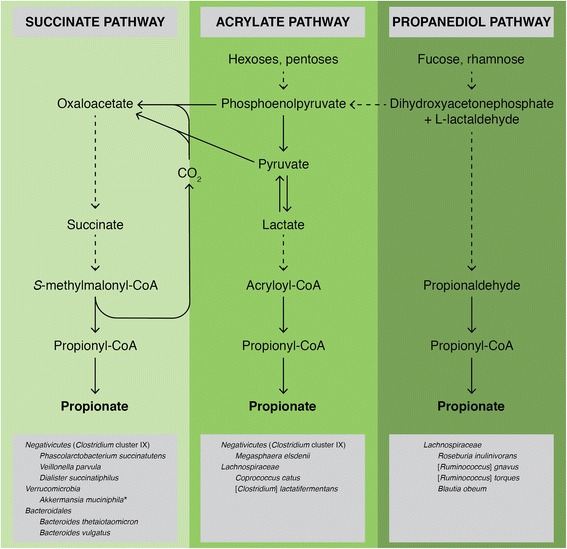
Background: Gut microbiota composition and function are symbiotically linked with host health and altered in metabolic, inflammatory and neurodegenerative disorders. Three recognised mechanisms exist by which the microbiome influences the gut-brain axis: modification of autonomic/sensorimotor connections, immune activation, and neuroendocrine pathway regulation. We hypothesised interactions between circulating gut-derived microbial metabolites, and the blood-brain barrier (BBB) also contribute to the gut-brain axis. Propionate, produced from dietary substrates by colonic bacteria, stimulates intestinal gluconeogenesis and is associated with reduced stress behaviours, but its potential endocrine role has not been addressed.
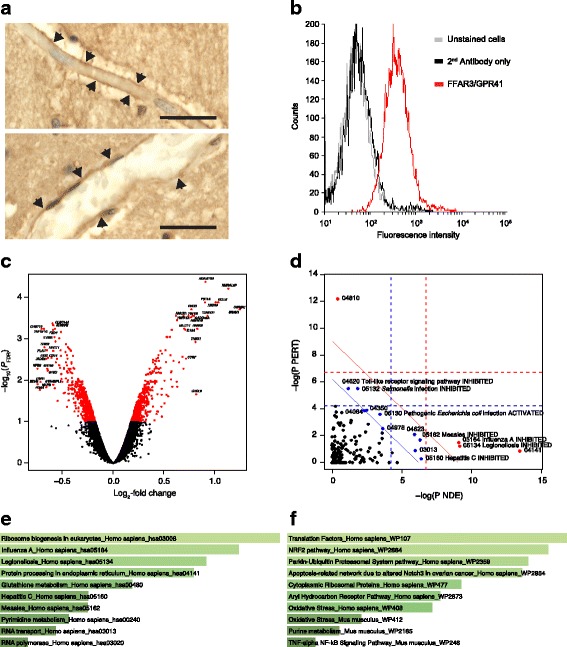
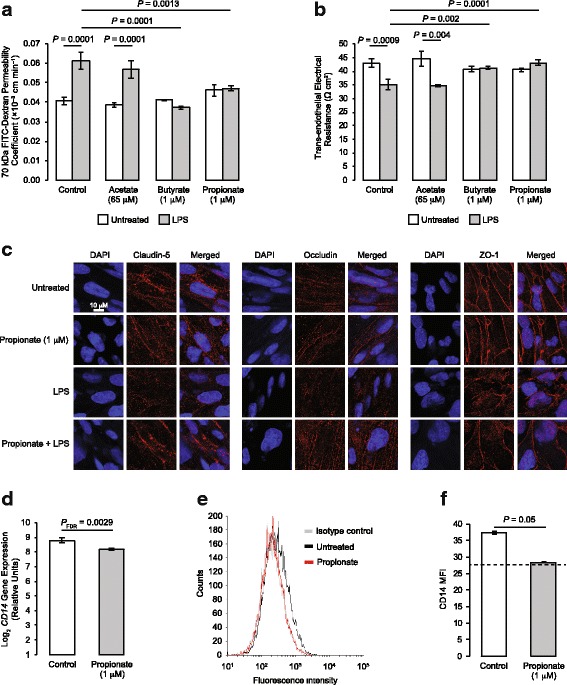
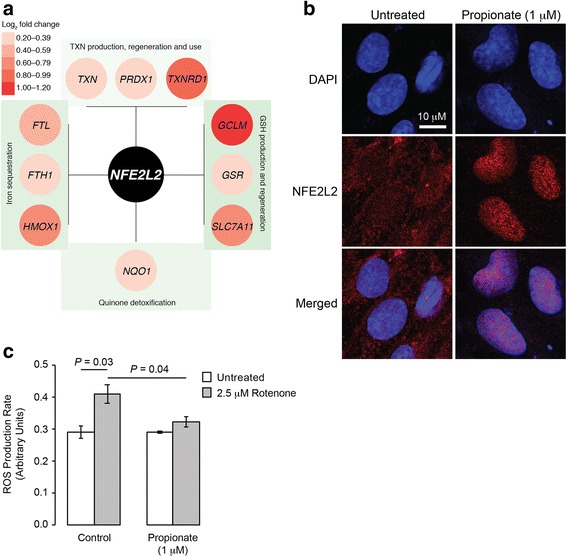
Results: After demonstrating expression of the propionate receptor FFAR3 on human brain endothelium, we examined the impact of a physiologically relevant propionate concentration (1 μM) on BBB properties in vitro. Propionate inhibited pathways associated with non-specific microbial infections via a CD14-dependent mechanism, suppressed expression of LRP-1 and protected the BBB from oxidative stress via NRF2 (NFE2L2) signalling.
Conclusions: Together, these results suggest gut-derived microbial metabolites interact with the BBB, representing a fourth facet of the gut-brain axis that warrants further attention.
Conflict of interest statement
Ethics approval and consent to participate
Not applicable
Consent for publication
Not applicable
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
