

Protein S-Nitrosylation Controls Glycogen Synthase Kinase 3β Function Independent of Its Phosphorylation State
- PMID: 29563102
- PMCID: PMC6047766
- DOI: 10.1161/CIRCRESAHA.118.312789
Protein S-Nitrosylation Controls Glycogen Synthase Kinase 3β Function Independent of Its Phosphorylation State
Abstract
Rationale: GSK-3β (glycogen synthase kinase 3β) is a multifunctional and constitutively active kinase known to regulate a myriad of cellular processes. The primary mechanism to regulate its function is through phosphorylation-dependent inhibition at serine-9 residue. Emerging evidence indicates that there may be alternative mechanisms that control GSK-3β for certain functions.
Objectives: Here, we sought to understand the role of protein S-nitrosylation (SNO) on the function of GSK-3β. SNO-dependent modulation of the localization of GSK-3β and its ability to phosphorylate downstream targets was investigated in vitro, and the network of proteins differentially impacted by phospho- or SNO-dependent GSK-3β regulation and in vivo SNO modification of key signaling kinases during the development of heart failure was also studied.
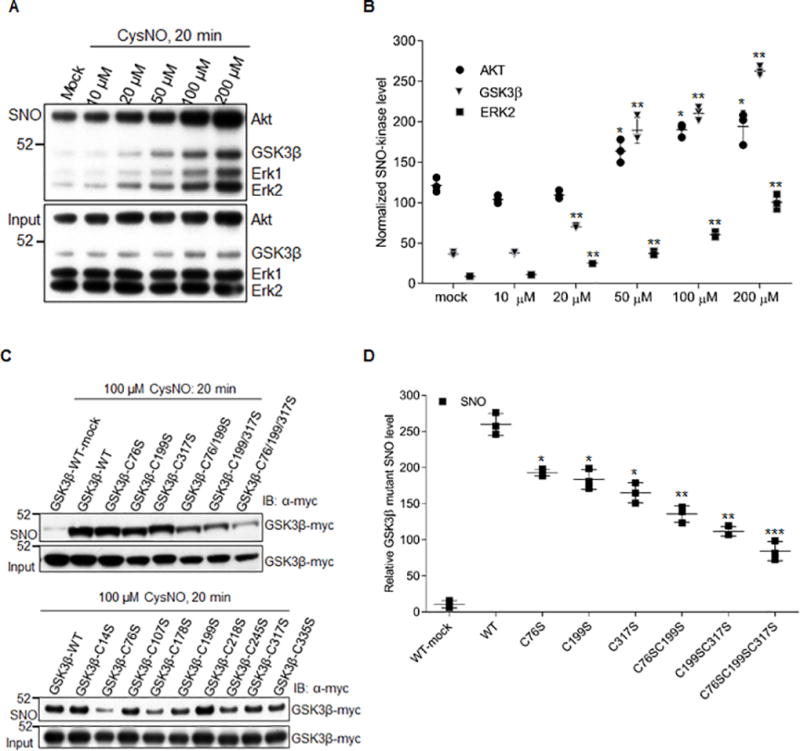
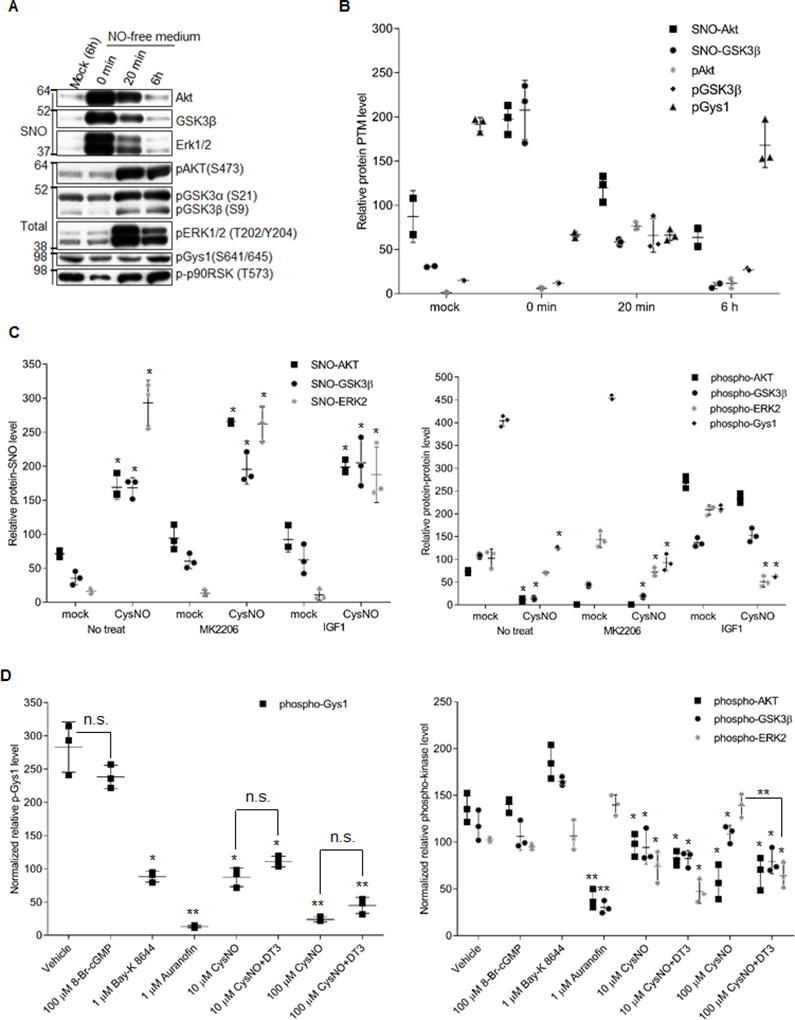
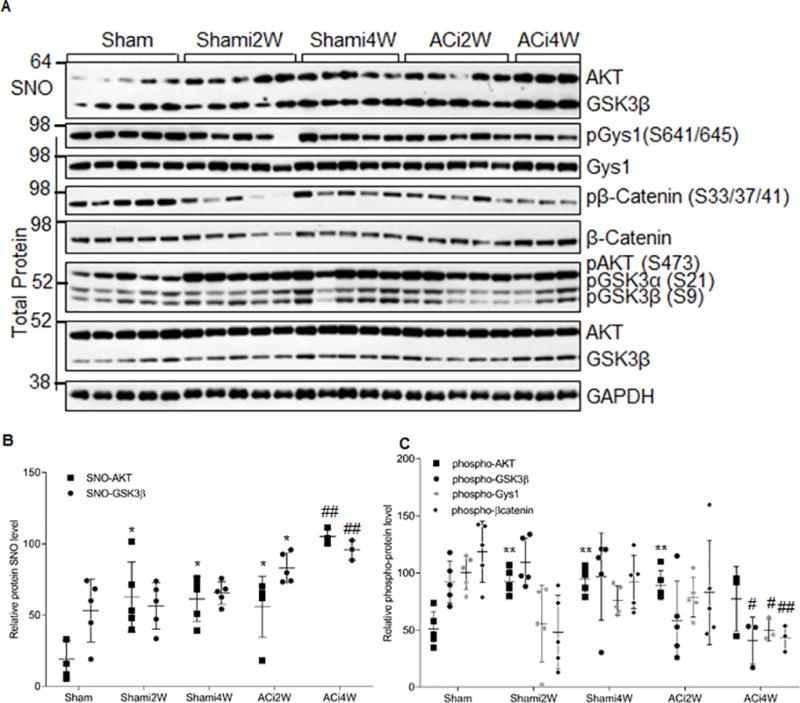
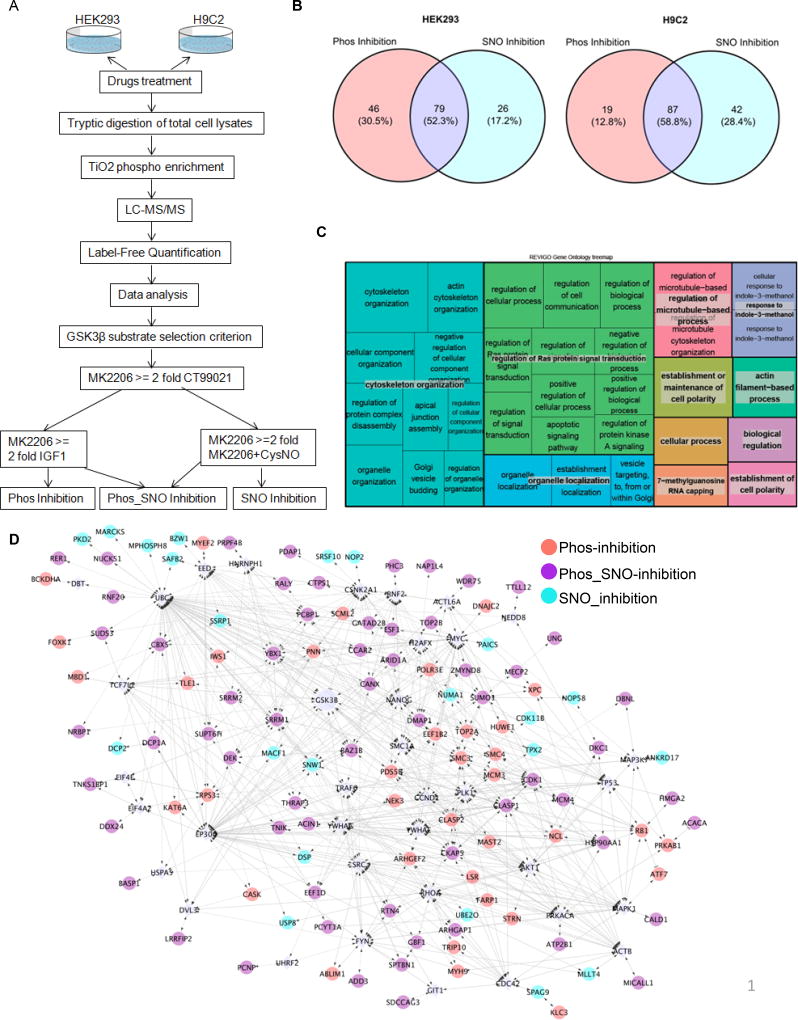
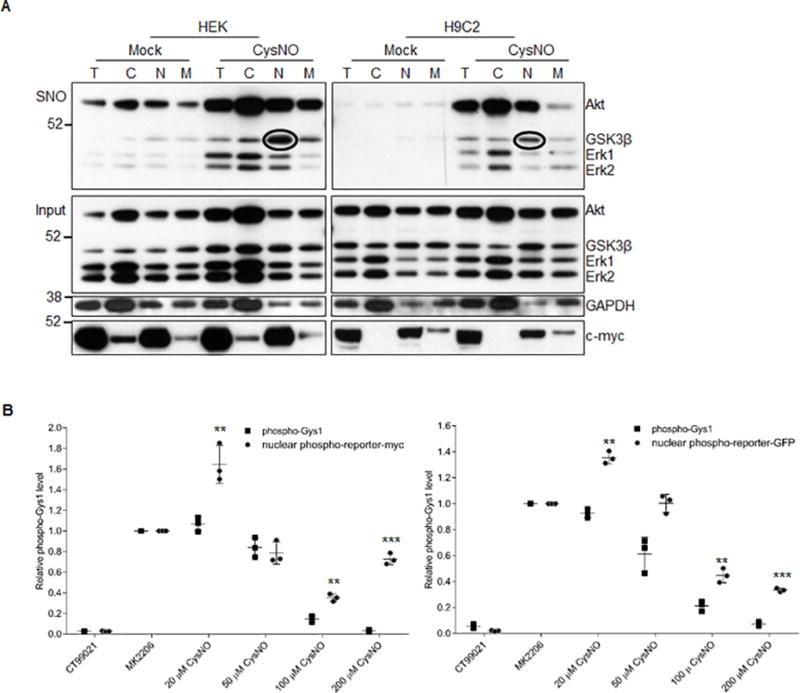
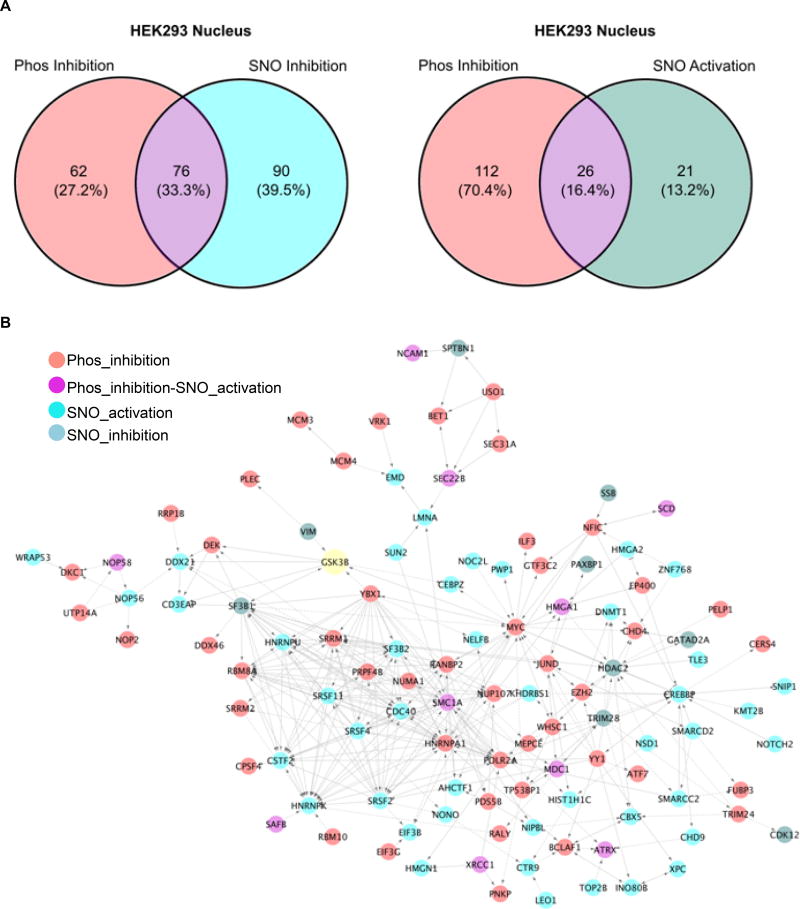
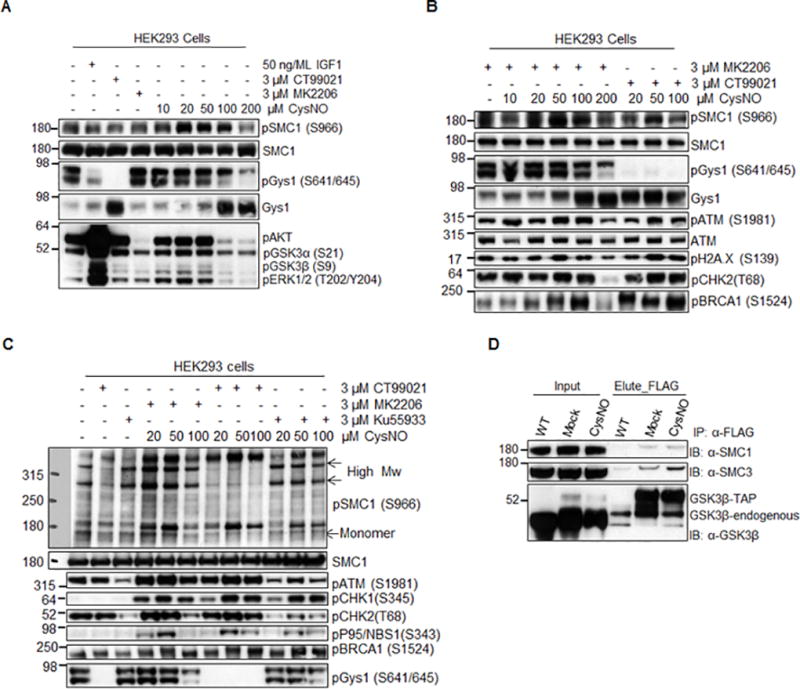
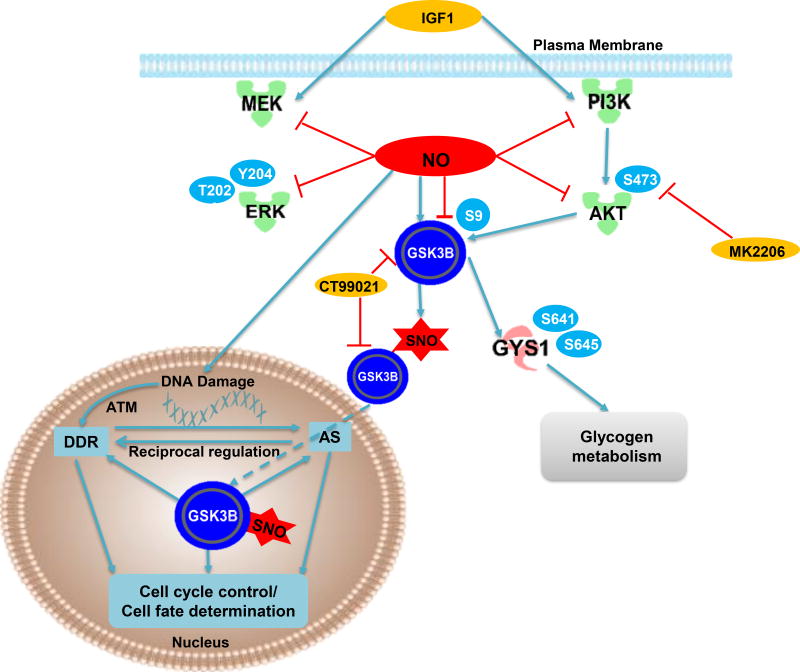
Methods and results: We found that GSK-3β undergoes site-specific SNO both in vitro, in HEK293 cells, H9C2 myoblasts, and primary neonatal rat ventricular myocytes, as well as in vivo, in hearts from an animal model of heart failure and sudden cardiac death. S-nitrosylation of GSK-3β significantly inhibits its kinase activity independent of the canonical phospho-inhibition pathway. S-nitrosylation of GSK-3β promotes its nuclear translocation and access to novel downstream phosphosubstrates which are enriched for a novel amino acid consensus sequence motif. Quantitative phosphoproteomics pathway analysis reveals that nuclear GSK-3β plays a central role in cell cycle control, RNA splicing, and DNA damage response.
Conclusions: The results indicate that SNO has a differential effect on the location and activity of GSK-3β in the cytoplasm versus the nucleus. SNO modification of GSK-3β occurs in vivo and could contribute to the pathobiology of heart failure and sudden cardiac death.
Keywords: S-nitrosylation; glycogen synthase kinase 3 beta; kinase-substrates interactome; nuclear translocation; redox regulation.
© 2018 American Heart Association, Inc.
Figures
Comment in
-
Cross Talk Between S-Nitrosylation and Phosphorylation Involving Kinases and Nitrosylases.Circ Res. 2018 May 25;122(11):1485-1487. doi: 10.1161/CIRCRESAHA.118.313109. Circ Res. 2018. PMID: 29798895 Free PMC article. No abstract available.
Similar articles
-
The mood stabilizing properties of AF3581, a novel potent GSK-3β inhibitor.Biomed Pharmacother. 2020 Aug;128:110249. doi: 10.1016/j.biopha.2020.110249. Epub 2020 May 26. Biomed Pharmacother. 2020. PMID: 32470749
-
GSK-3β promotes PA-induced apoptosis through changing β-arrestin 2 nucleus location in H9c2 cardiomyocytes.Apoptosis. 2016 Sep;21(9):1045-55. doi: 10.1007/s10495-016-1272-z. Apoptosis. 2016. PMID: 27431999
-
S-nitrosylation of Hsp90 promotes cardiac hypertrophy in mice through GSK3β signaling.Acta Pharmacol Sin. 2022 Aug;43(8):1979-1988. doi: 10.1038/s41401-021-00828-9. Epub 2021 Dec 21. Acta Pharmacol Sin. 2022. PMID: 34934196 Free PMC article.
-
Glycogen synthase kinase-3beta: a novel regulator of cardiac hypertrophy and development.Circ Res. 2002 May 31;90(10):1055-63. doi: 10.1161/01.res.0000018952.70505.f1. Circ Res. 2002. PMID: 12039794 Review.
-
Glycogen synthase kinase 3β in tumorigenesis and oncotherapy (Review).Oncol Rep. 2020 Dec;44(6):2373-2385. doi: 10.3892/or.2020.7817. Epub 2020 Oct 20. Oncol Rep. 2020. PMID: 33125126 Free PMC article. Review.
Cited by
-
The Role of Glycogen Synthase Kinase-3 in the Regulation of Ribosome Biogenesis in Rat Soleus Muscle under Disuse Conditions.Int J Mol Sci. 2022 Mar 2;23(5):2751. doi: 10.3390/ijms23052751. Int J Mol Sci. 2022. PMID: 35269893 Free PMC article.
-
Blocking Nitrosylation Induces Immunogenic Cell Death by Sensitizing NRAS-Mutant Melanoma to MEK Inhibitors.Cancer Res. 2025 Jun 16;85(12):2268-2287. doi: 10.1158/0008-5472.CAN-24-0693. Cancer Res. 2025. PMID: 40287947 Free PMC article.
-
Integrated Dissection of Cysteine Oxidative Post-translational Modification Proteome During Cardiac Hypertrophy.J Proteome Res. 2018 Dec 7;17(12):4243-4257. doi: 10.1021/acs.jproteome.8b00372. Epub 2018 Aug 31. J Proteome Res. 2018. PMID: 30141336 Free PMC article.
-
Effects of Gestational Hypoxia on PGC1α and Mitochondrial Acetylation in Fetal Guinea Pig Hearts.Reprod Sci. 2023 Oct;30(10):2996-3009. doi: 10.1007/s43032-023-01245-5. Epub 2023 May 3. Reprod Sci. 2023. PMID: 37138147 Free PMC article.
-
Regulation of Three Key Kinases of Brassinosteroid Signaling Pathway.Int J Mol Sci. 2020 Jun 18;21(12):4340. doi: 10.3390/ijms21124340. Int J Mol Sci. 2020. PMID: 32570783 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
