

EB1 binding restricts STIM1 translocation to ER-PM junctions and regulates store-operated Ca2+ entry
- PMID: 29563214
- PMCID: PMC5987725
- DOI: 10.1083/jcb.201711151
EB1 binding restricts STIM1 translocation to ER-PM junctions and regulates store-operated Ca2+ entry
Abstract
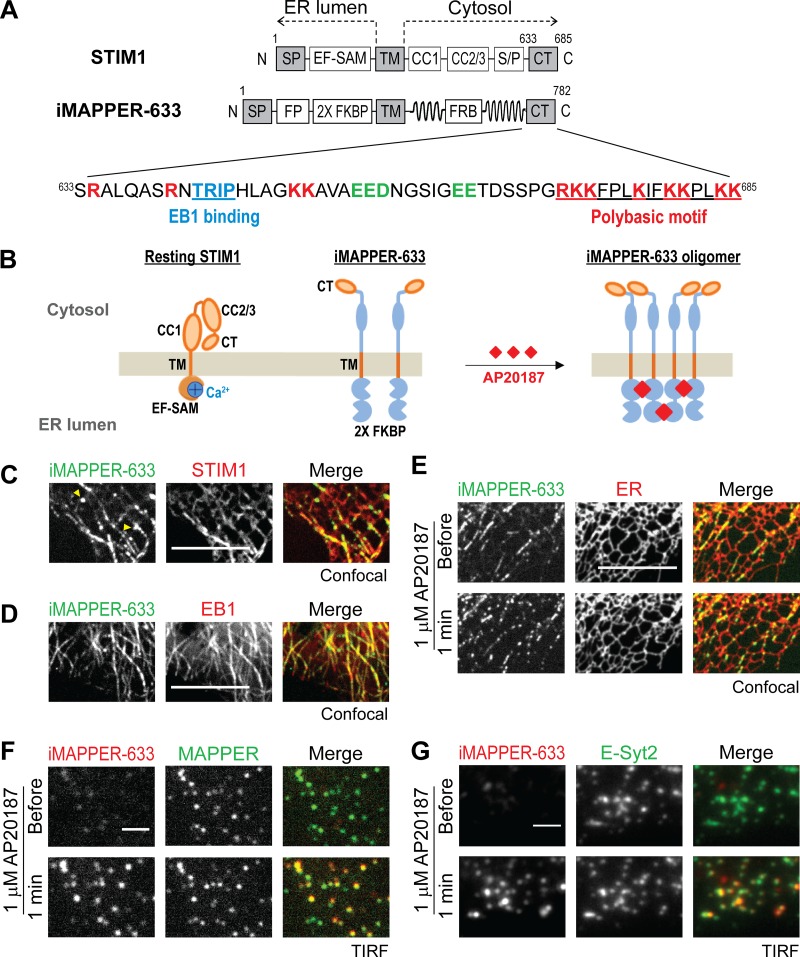
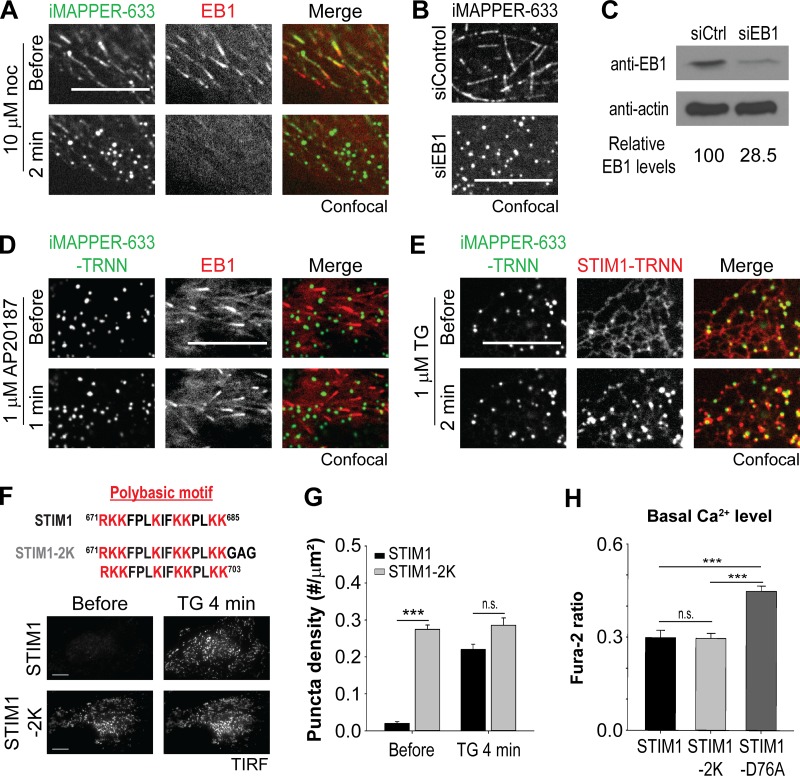
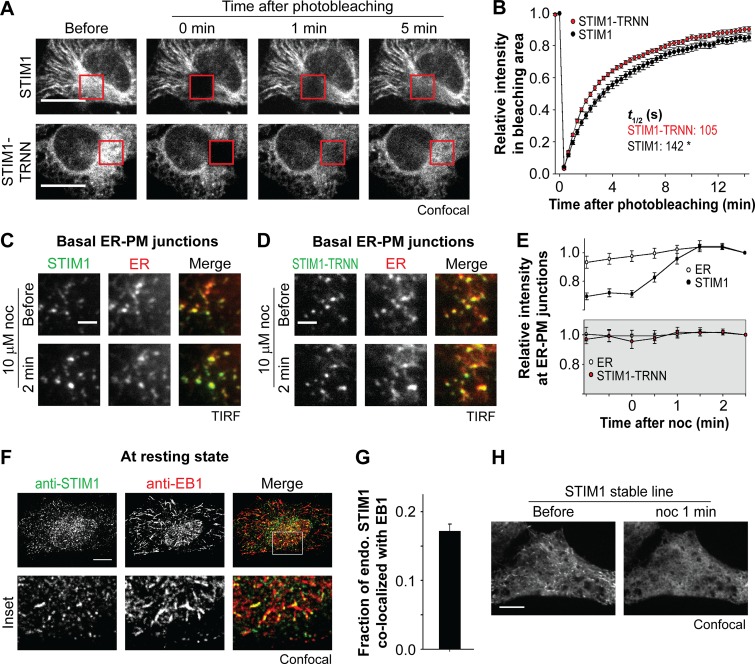
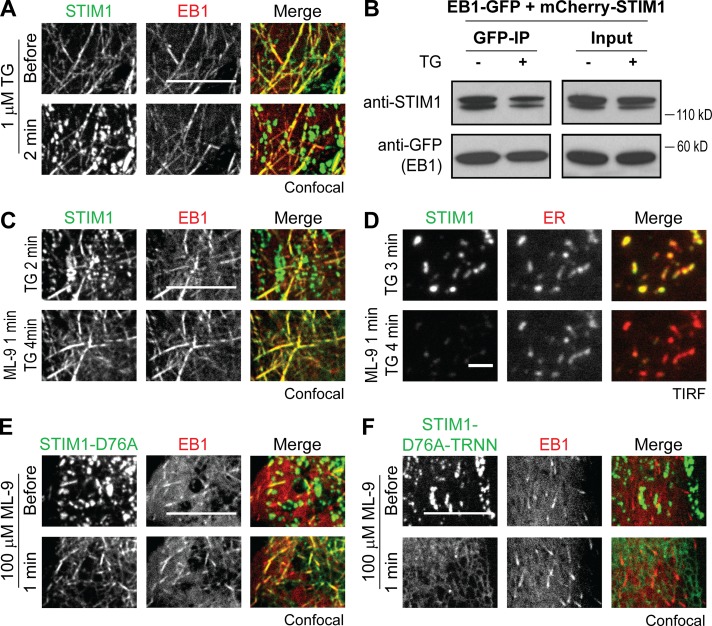
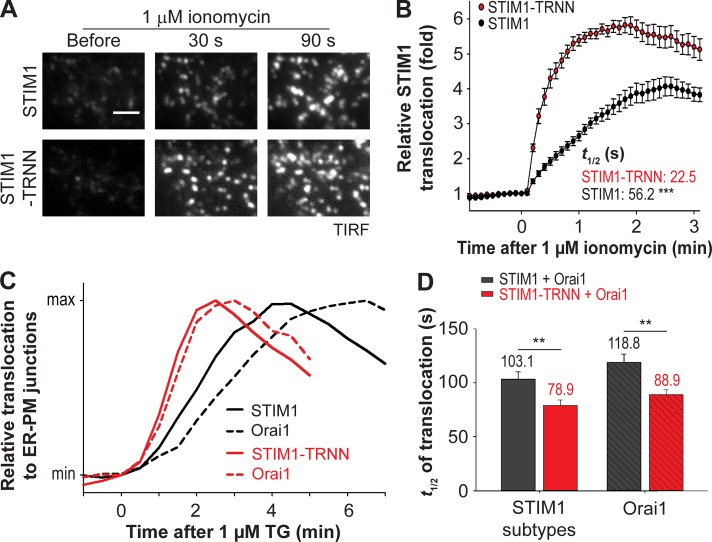
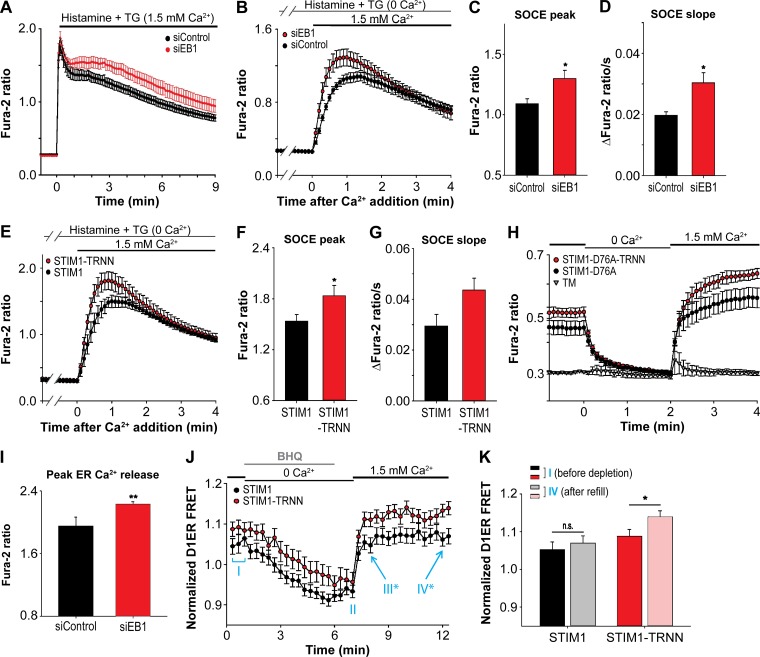
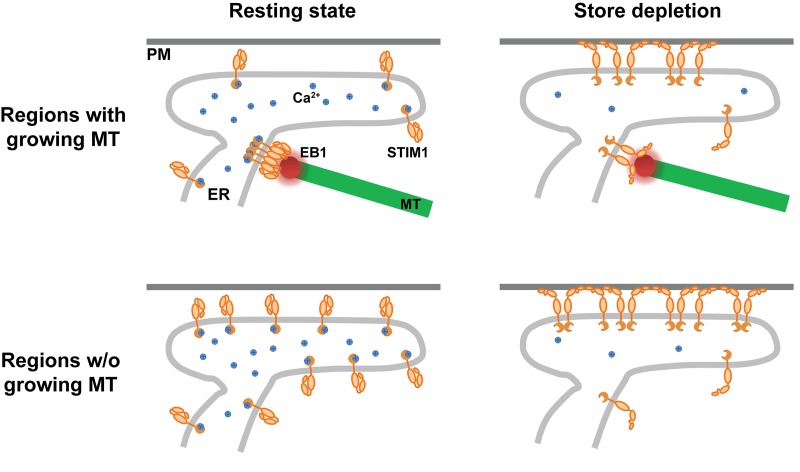
The endoplasmic reticulum (ER) Ca2+ sensor STIM1 forms oligomers and translocates to ER-plasma membrane (PM) junctions to activate store-operated Ca2+ entry (SOCE) after ER Ca2+ depletion. STIM1 also interacts with EB1 and dynamically tracks microtubule (MT) plus ends. Nevertheless, the role of STIM1-EB1 interaction in regulating SOCE remains unresolved. Using live-cell imaging combined with a synthetic construct approach, we found that EB1 binding constitutes a trapping mechanism restricting STIM1 targeting to ER-PM junctions. We further showed that STIM1 oligomers retain EB1 binding ability in ER Ca2+-depleted cells. By trapping STIM1 molecules at dynamic contacts between the ER and MT plus ends, EB1 binding delayed STIM1 translocation to ER-PM junctions during ER Ca2+ depletion and prevented excess SOCE and ER Ca2+ overload. Our study suggests that STIM1-EB1 interaction shapes the kinetics and amplitude of local SOCE in cellular regions with growing MTs and contributes to spatiotemporal regulation of Ca2+ signaling crucial for cellular functions and homeostasis.
© 2018 Chang et al.
Figures
References
-
- Baba Y., Hayashi K., Fujii Y., Mizushima A., Watarai H., Wakamori M., Numaga T., Mori Y., Iino M., Hikida M., and Kurosaki T.. 2006. Coupling of STIM1 to store-operated Ca2+ entry through its constitutive and inducible movement in the endoplasmic reticulum. Proc. Natl. Acad. Sci. USA. 103:16704–16709. 10.1073/pnas.0608358103 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
