

Activation and Induction of Antigen-Specific T Follicular Helper Cells Play a Critical Role in Live-Attenuated Influenza Vaccine-Induced Human Mucosal Anti-influenza Antibody Response
- PMID: 29563292
- PMCID: PMC5952133
- DOI: 10.1128/JVI.00114-18
Activation and Induction of Antigen-Specific T Follicular Helper Cells Play a Critical Role in Live-Attenuated Influenza Vaccine-Induced Human Mucosal Anti-influenza Antibody Response
Abstract
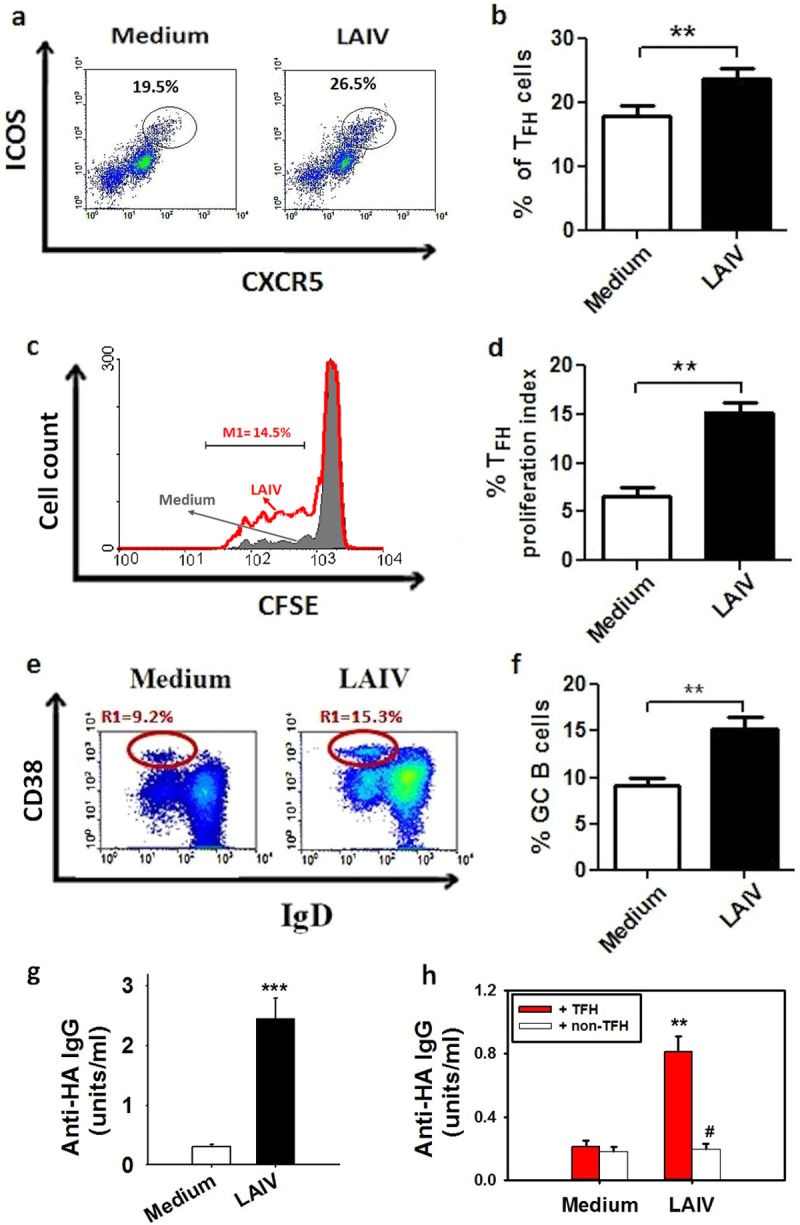
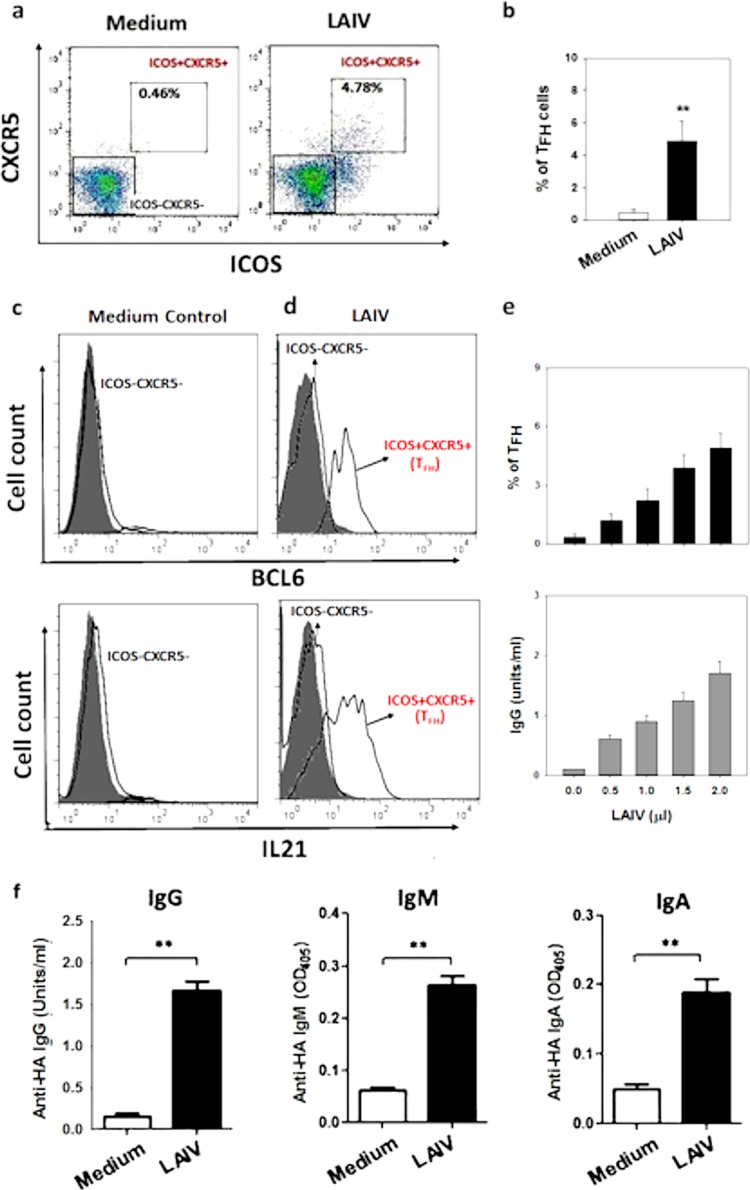
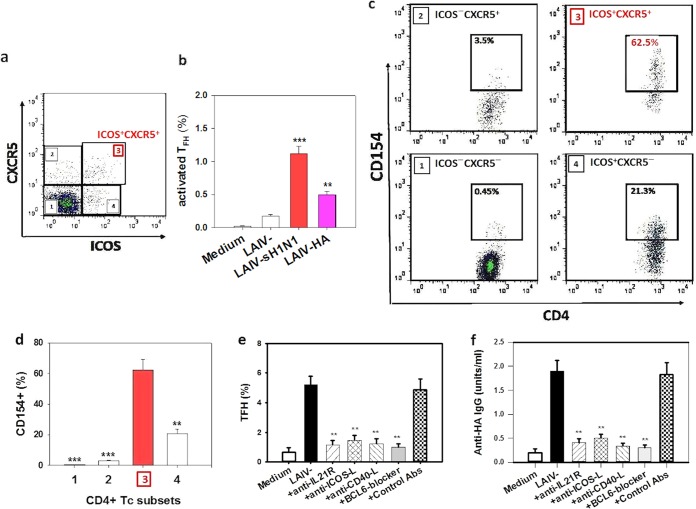
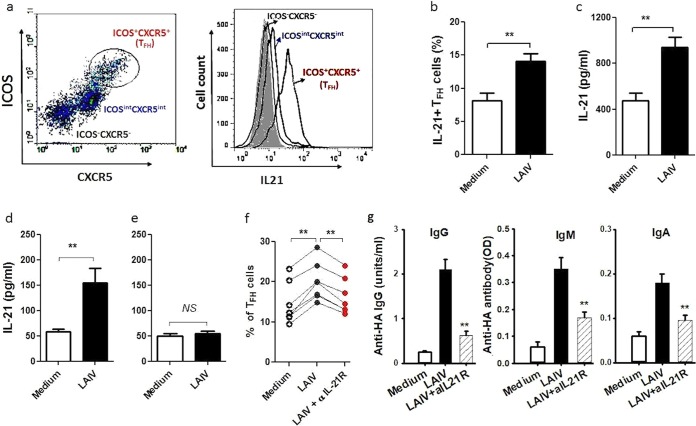
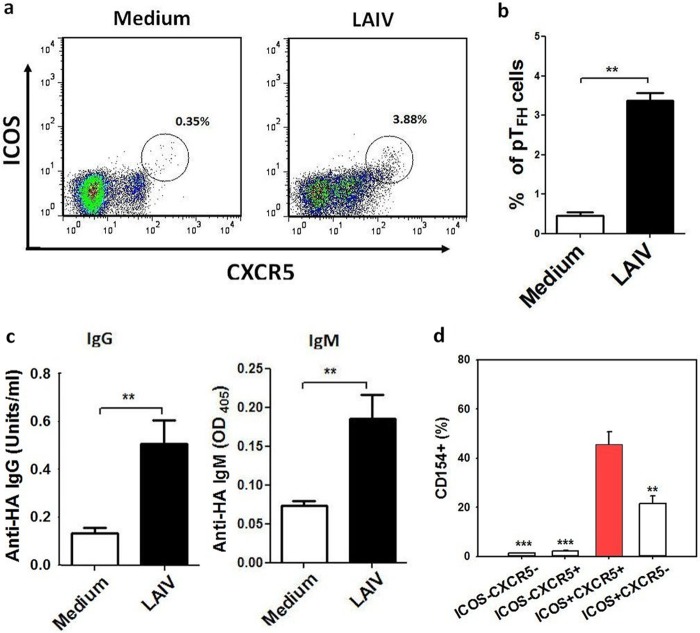
There is increasing interest recently in developing intranasal vaccines against respiratory tract infections. The antibody response is critical for vaccine-induced protection, and T follicular helper cells (TFH) are considered important for mediating the antibody response. Most data supporting the role for TFH in the antibody response are from animal studies, and direct evidence from humans is limited, apart from the presence of TFH-like cells in blood. We studied the activation and induction of TFH and their role in the anti-influenza antibody response induced by a live-attenuated influenza vaccine (LAIV) in human nasopharynx-associated lymphoid tissue (NALT). TFH activation in adenotonsillar tissues was analyzed by flow cytometry, and anti-hemagglutinin (anti-HA) antibodies were examined following LAIV stimulation of tonsillar mononuclear cells (MNC). Induction of antigen-specific TFH by LAIV was studied by flow cytometry analysis of induced TFH and CD154 expression. LAIV induced TFH proliferation, which correlated with anti-HA antibody production, and TFH were shown to be critical for the antibody response. Induction of TFH from naive T cells by LAIV was shown in newly induced TFH expressing BCL6 and CD21, followed by the detection of anti-HA antibodies. Antigen specificity of LAIV-induced TFH was demonstrated by expression of the antigen-specific T cell activation marker CD154 upon challenge by H1N1 virus antigen or HA. LAIV-induced TFH differentiation was inhibited by BCL6, interleukin-21 (IL-21), ICOS, and CD40 signaling blocking, and that diminished anti-HA antibody production. In conclusion, we demonstrated the induction by LAIV of antigen-specific TFH in human NALT that provide critical support for the anti-influenza antibody response. Promoting antigen-specific TFH in NALT by use of intranasal vaccines may provide an effective vaccination strategy against respiratory infections in humans.IMPORTANCE Airway infections, such as influenza, are common in humans. Intranasal vaccination has been considered a biologically relevant and effective way of immunization against airway infection. The vaccine-induced antibody response is crucial for protection against infection. Recent data from animal studies suggest that one type of T cells, TFH, are important for the antibody response. However, data on whether TFH-mediated help for antibody production operates in humans are limited due to the lack of access to human immune tissue containing TFH In this study, we demonstrate the induction of TFH in human immune tissue, providing critical support for the anti-influenza antibody response, by use of an intranasal influenza vaccine. Our findings provide direct evidence that TFH play a critical role in vaccine-induced immunity in humans and suggest a novel strategy for promoting such cells by use of intranasal vaccines against respiratory infections.
Keywords: LAIV; NALT; T follicular helper cell; TFH; antibody response; influenza vaccine; mucosal immunity; nasopharynx-associated lymphoid tissue.
Copyright © 2018 Aljurayyan et al.
Figures
References
-
- Slight SR, Rangel-Moreno J, Gopal R, Lin Y, Fallert Junecko BA, Mehra S, Selman M, Becerril-Villanueva E, Baquera-Heredia J, Pavon L, Kaushal D, Reinhart TA, Randall TD, Khader SA. 2013. CXCR5+ T helper cells mediate protective immunity against tuberculosis. J Clin Invest 123:712–726. doi: 10.1172/JCI65728. - DOI - PMC - PubMed
-
- Herati RS, Reuter MA, Dolfi DV, Mansfield KD, Aung H, Badwan OZ, Kurupati RK, Kannan S, Ertl H, Schmader KE, Betts MR, Canaday DH, Wherry EJ. 2014. Circulating CXCR5(+)PD-1(+) response predicts influenza vaccine antibody responses in young adults but not elderly adults. J Immunol 193:3528–3537. doi: 10.4049/jimmunol.1302503. - DOI - PMC - PubMed
-
- Morita R, Schmitt N, Bentebibel S-E, Ranganathan R, Bourdery L, Zurawski G, Foucat E, Dullaers M, Oh S, Sabzghabaei N, Lavecchio EM, Punaro M, Pascual V, Banchereau J, Ueno H. 2011. Human blood CXCR5(+)CD4(+) T cells are counterparts of T follicular cells and contain specific subsets that differentially support antibody secretion. Immunity 34:108–121. doi: 10.1016/j.immuni.2010.12.012. - DOI - PMC - PubMed
-
- Bentebibel S-E, Lopez S, Obermoser G, Schmitt N, Mueller C, Harrod C, Flano E, Mejias A, Albrecht RA, Blankenship D, Xu H, Pascual V, Banchereau J, Garcia-Sastre A, Palucka AK, Ramilo O, Ueno H. 2013. Induction of ICOS+CXCR3+CXCR5+ TH cells correlates with antibody responses to influenza vaccination. Sci Transl Med 5:176ra132. doi: 10.1126/scitranslmed.3005191. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
