

The Involvement of Endogenous Neural Oscillations in the Processing of Rhythmic Input: More Than a Regular Repetition of Evoked Neural Responses
- PMID: 29563860
- PMCID: PMC5845906
- DOI: 10.3389/fnins.2018.00095
The Involvement of Endogenous Neural Oscillations in the Processing of Rhythmic Input: More Than a Regular Repetition of Evoked Neural Responses
Abstract
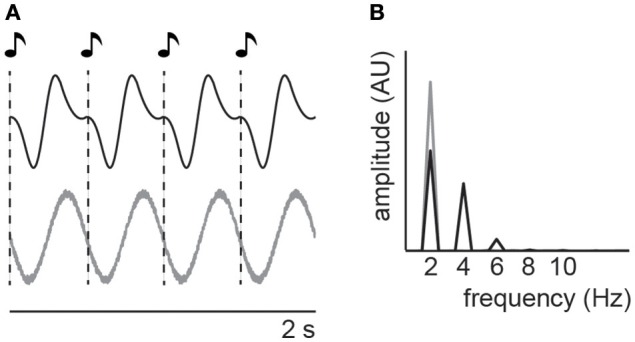
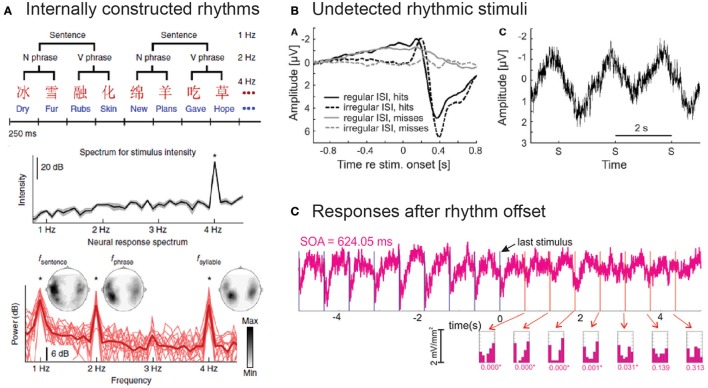
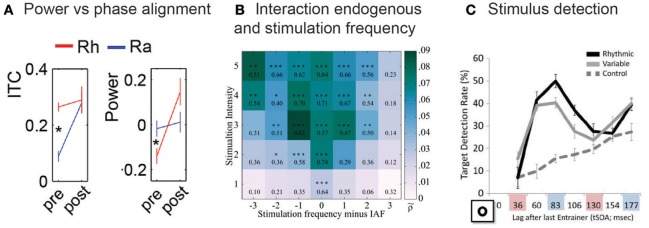
It is undisputed that presenting a rhythmic stimulus leads to a measurable brain response that follows the rhythmic structure of this stimulus. What is still debated, however, is the question whether this brain response exclusively reflects a regular repetition of evoked responses, or whether it also includes entrained oscillatory activity. Here we systematically present evidence in favor of an involvement of entrained neural oscillations in the processing of rhythmic input while critically pointing out which questions still need to be addressed before this evidence could be considered conclusive. In this context, we also explicitly discuss the potential functional role of such entrained oscillations, suggesting that these stimulus-aligned oscillations reflect, and serve as, predictive processes, an idea often only implicitly assumed in the literature.
Keywords: ERP; endogenous; entrainment; evoked response; oscillation; phase; power.
Figures
Similar articles
-
Sustained neural rhythms reveal endogenous oscillations supporting speech perception.PLoS Biol. 2021 Feb 26;19(2):e3001142. doi: 10.1371/journal.pbio.3001142. eCollection 2021 Feb. PLoS Biol. 2021. PMID: 33635855 Free PMC article.
-
Phase Alignment of Low-Frequency Neural Activity to the Amplitude Envelope of Speech Reflects Evoked Responses to Acoustic Edges, Not Oscillatory Entrainment.J Neurosci. 2023 May 24;43(21):3909-3921. doi: 10.1523/JNEUROSCI.1663-22.2023. Epub 2023 Apr 26. J Neurosci. 2023. PMID: 37185238 Free PMC article.
-
Neural Entrainment in Drum Rhythms with Silent Breaks: Evidence from Steady-state Evoked and Event-related Potentials.J Cogn Neurosci. 2016 Dec;28(12):1865-1877. doi: 10.1162/jocn_a_01013. Epub 2016 Jul 26. J Cogn Neurosci. 2016. PMID: 27458750
-
Oscillations in the auditory system and their possible role.Neurosci Biobehav Rev. 2020 Jun;113:507-528. doi: 10.1016/j.neubiorev.2020.03.030. Epub 2020 Apr 13. Neurosci Biobehav Rev. 2020. PMID: 32298712 Review.
-
Oscillatory Mechanisms of Stimulus Processing and Selection in the Visual and Auditory Systems: State-of-the-Art, Speculations and Suggestions.Front Neurosci. 2017 May 26;11:296. doi: 10.3389/fnins.2017.00296. eCollection 2017. Front Neurosci. 2017. PMID: 28603483 Free PMC article. Review.
Cited by
-
Hearing Impaired Participants Improve More Under Envelope-Transcranial Alternating Current Stimulation When Signal to Noise Ratio Is High.Neurosci Insights. 2021 Feb 24;16:2633105520988854. doi: 10.1177/2633105520988854. eCollection 2021. Neurosci Insights. 2021. PMID: 33709079 Free PMC article.
-
Distinct roles of delta- and theta-band neural tracking for sharpening and predictive coding of multi-level speech features during spoken language processing.Hum Brain Mapp. 2023 Dec 1;44(17):6149-6172. doi: 10.1002/hbm.26503. Epub 2023 Oct 11. Hum Brain Mapp. 2023. PMID: 37818940 Free PMC article.
-
Processing rhythm in speech and music: Shared mechanisms and implications for developmental speech and language disorders.Neuropsychology. 2021 Nov;35(8):771-791. doi: 10.1037/neu0000766. Epub 2021 Aug 26. Neuropsychology. 2021. PMID: 34435803 Free PMC article. Review.
-
Double dissociation of single-interval and rhythmic temporal prediction in cerebellar degeneration and Parkinson's disease.Proc Natl Acad Sci U S A. 2018 Nov 27;115(48):12283-12288. doi: 10.1073/pnas.1810596115. Epub 2018 Nov 13. Proc Natl Acad Sci U S A. 2018. PMID: 30425170 Free PMC article.
-
Short-Term Effect of Auditory Stimulation on Neural Activities: A Scoping Review of Longitudinal Electroencephalography and Magnetoencephalography Studies.Brain Sci. 2024 Jan 26;14(2):131. doi: 10.3390/brainsci14020131. Brain Sci. 2024. PMID: 38391706 Free PMC article.
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources