

Ten Years of High Resolution Structural Research on the Voltage Dependent Anion Channel (VDAC)-Recent Developments and Future Directions
- PMID: 29563878
- PMCID: PMC5845903
- DOI: 10.3389/fphys.2018.00108
Ten Years of High Resolution Structural Research on the Voltage Dependent Anion Channel (VDAC)-Recent Developments and Future Directions
Abstract
Mitochondria are evolutionarily related to Gram-negative bacteria and both comprise two membrane systems with strongly differing protein composition. The major protein in the outer membrane of mitochondria is the voltage-dependent anion channel (VDAC), which mediates signal transmission across the outer membrane but also the exchange of metabolites, most importantly ADP and ATP. More than 30 years after its discovery three identical high-resolution structures were determined in 2008. These structures show a 19-stranded anti-parallel beta-barrel with an N-terminal helix located inside. An odd number of beta-strands is also shared by Tom40, another member of the VDAC superfamily. This indicates that this superfamily is evolutionarily relatively young and that it has emerged in the context of mitochondrial evolution. New structural information obtained during the last decade on Tom40 can be used to cross-validate the structure of VDAC and vice versa. Interpretation of biochemical and biophysical studies on both protein channels now rests on a solid basis of structural data. Over the past 10 years, complementary structural and functional information on proteins of the VDAC superfamily has been collected from in-organello, in-vitro, and in silico studies. Most of these findings have confirmed the validity of the original structures. This short article briefly reviews the most important advances on the structure and function of VDAC superfamily members collected during the last decade and summarizes how they enhanced our understanding of the channel.
Keywords: NMR; Tom40; VDAC; structural biology; x-ray.
Figures
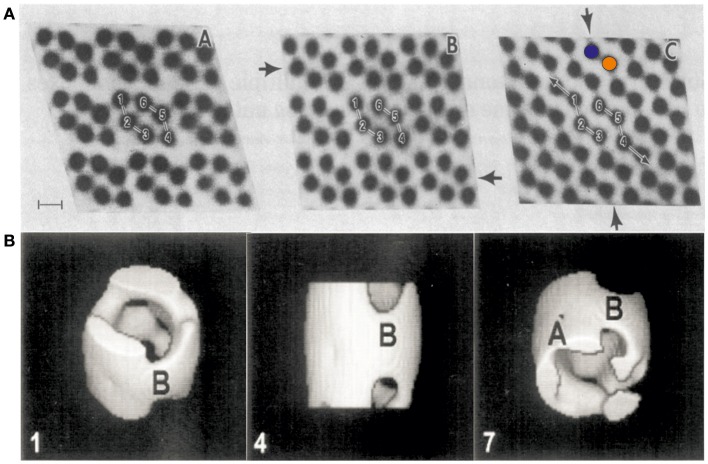
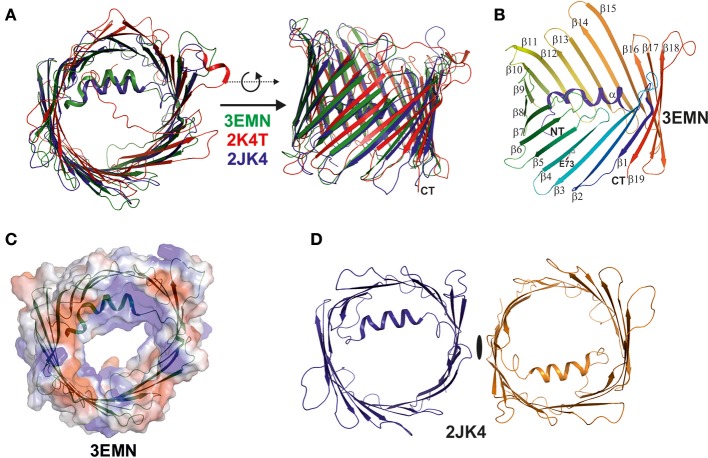
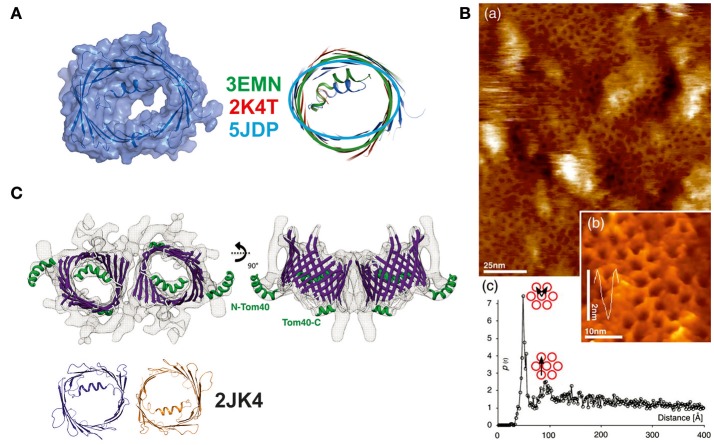
References
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous