

Generation of a Synthetic Human Chromosome with Two Centromeric Domains for Advanced Epigenetic Engineering Studies
- PMID: 29565577
- PMCID: PMC5951608
- DOI: 10.1021/acssynbio.8b00018
Generation of a Synthetic Human Chromosome with Two Centromeric Domains for Advanced Epigenetic Engineering Studies
Abstract
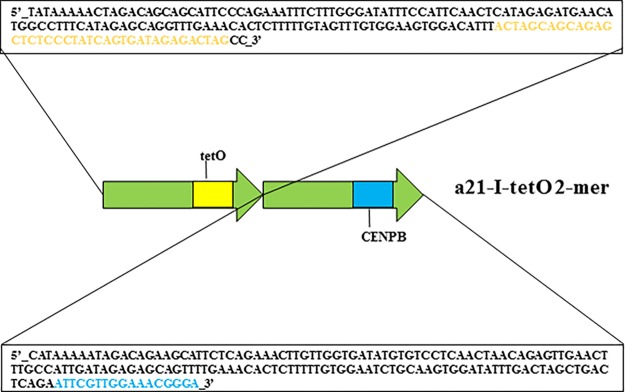
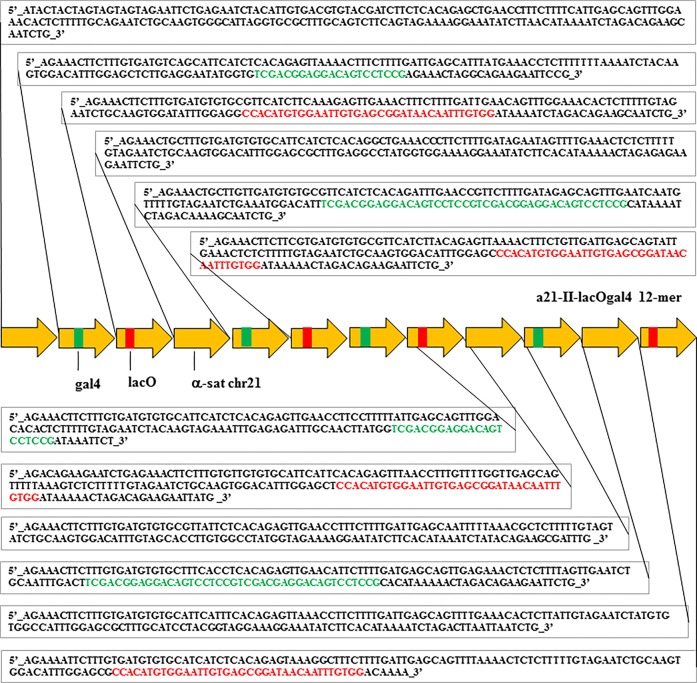
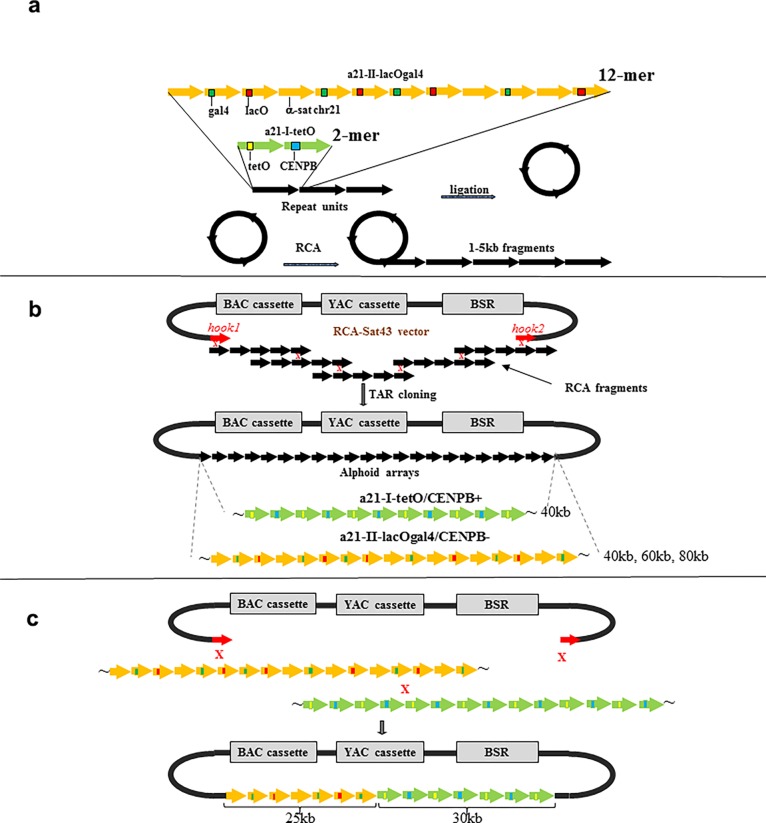
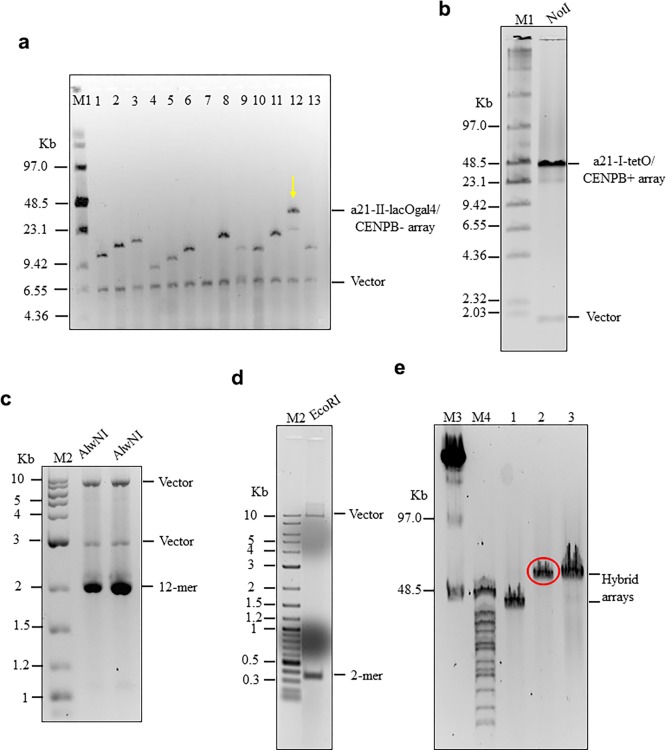
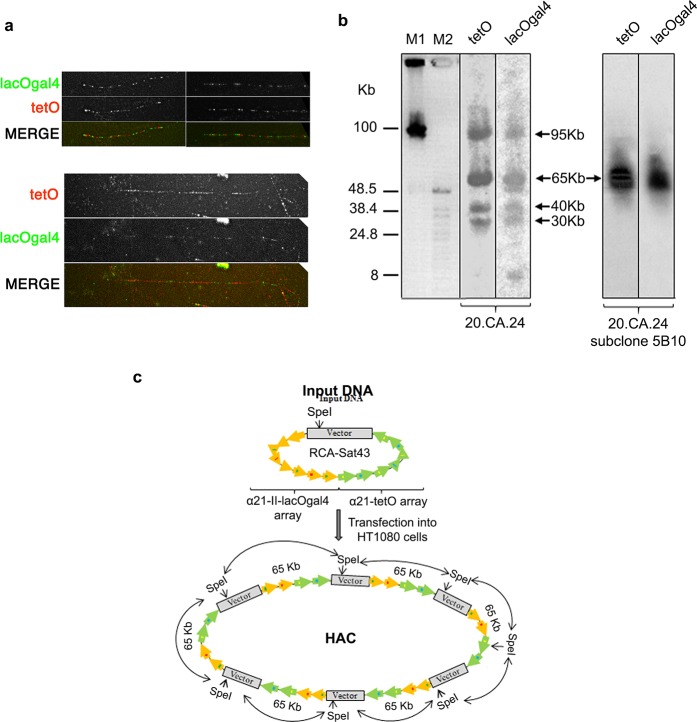
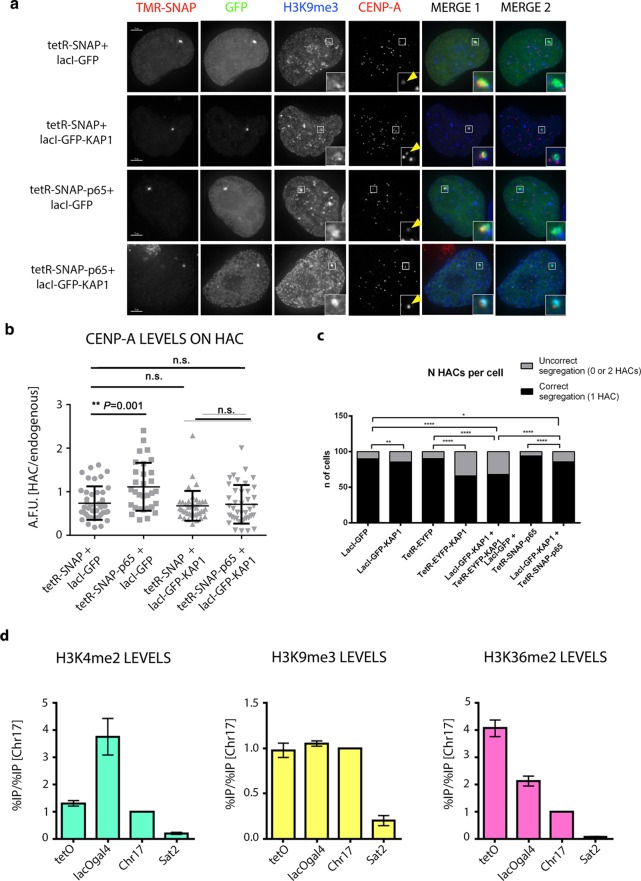
It is generally accepted that chromatin containing the histone H3 variant CENP-A is an epigenetic mark maintaining centromere identity. However, the pathways leading to the formation and maintenance of centromere chromatin remain poorly characterized due to difficulties of analysis of centromeric repeats in native chromosomes. To address this problem, in our previous studies we generated a human artificial chromosome (HAC) whose centromere contains a synthetic alpha-satellite (alphoid) DNA array containing the tetracycline operator, the alphoidtetO-HAC. The presence of tetO sequences allows the specific targeting of the centromeric region in the HAC with different chromatin modifiers fused to the tetracycline repressor. The alphoidtetO-HAC has been extensively used to investigate protein interactions within the kinetochore and to define the epigenetic signature of centromeric chromatin to maintain a functional kinetochore. In this study, we developed a novel synthetic HAC containing two alphoid DNA arrays with different targeting sequences, tetO, lacO and gal4, the alphoidhybrid-HAC. This new HAC can be used for detailed epigenetic engineering studies because its kinetochore can be simultaneously or independently targeted by different chromatin modifiers and other fusion proteins.
Keywords: centromere; chromosome segregation; human artificial chromosome; kinetochore; mitosis.
Conflict of interest statement
The authors declare no competing financial interest.
Figures


References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
