

The BRD3 ET domain recognizes a short peptide motif through a mechanism that is conserved across chromatin remodelers and transcriptional regulators
- PMID: 29567837
- PMCID: PMC5949996
- DOI: 10.1074/jbc.RA117.000678
The BRD3 ET domain recognizes a short peptide motif through a mechanism that is conserved across chromatin remodelers and transcriptional regulators
Abstract
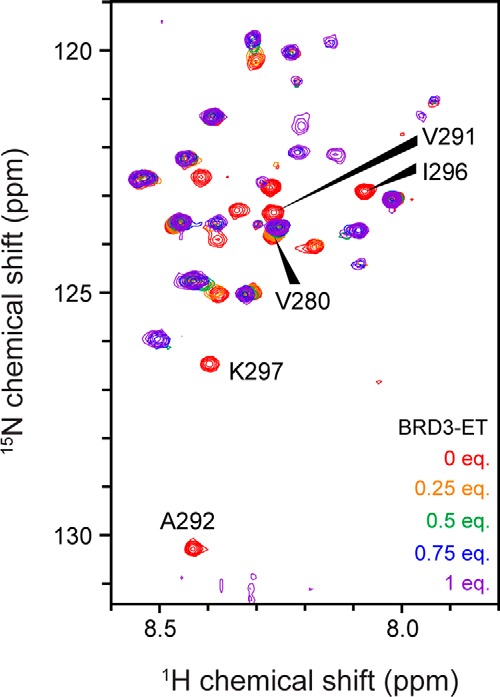
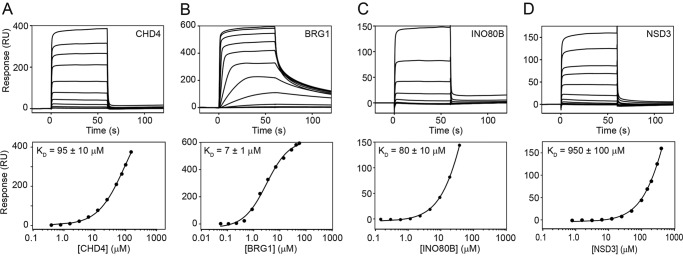
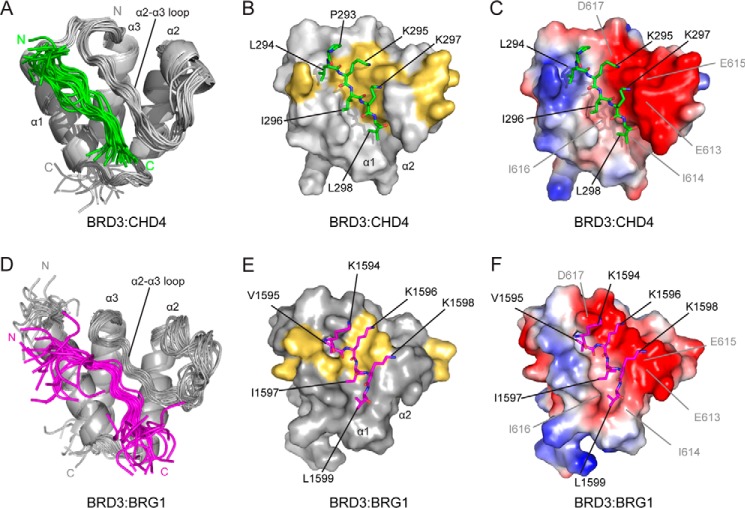
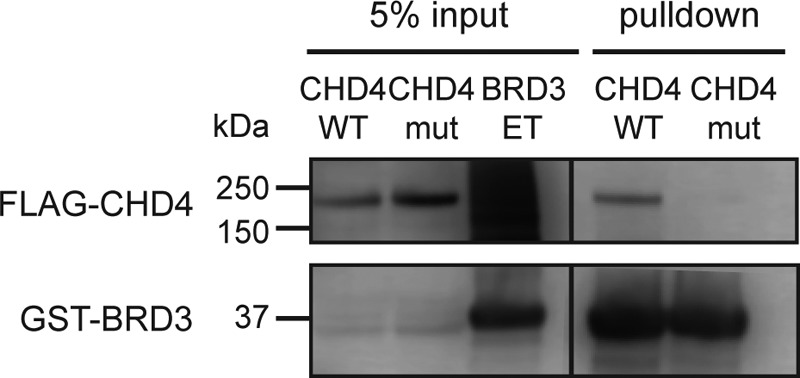
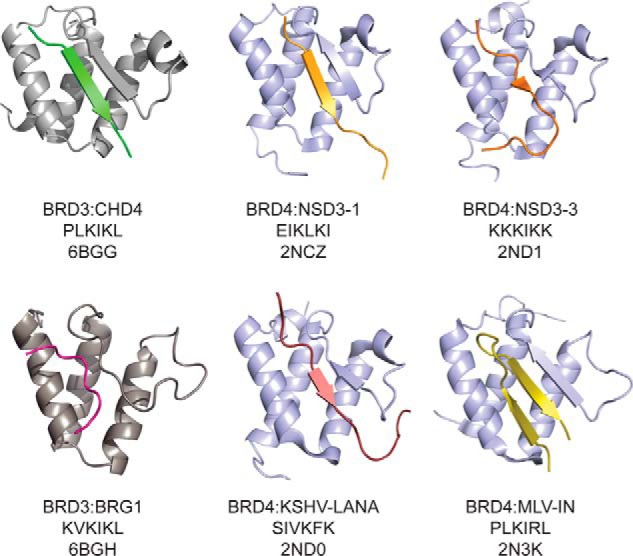
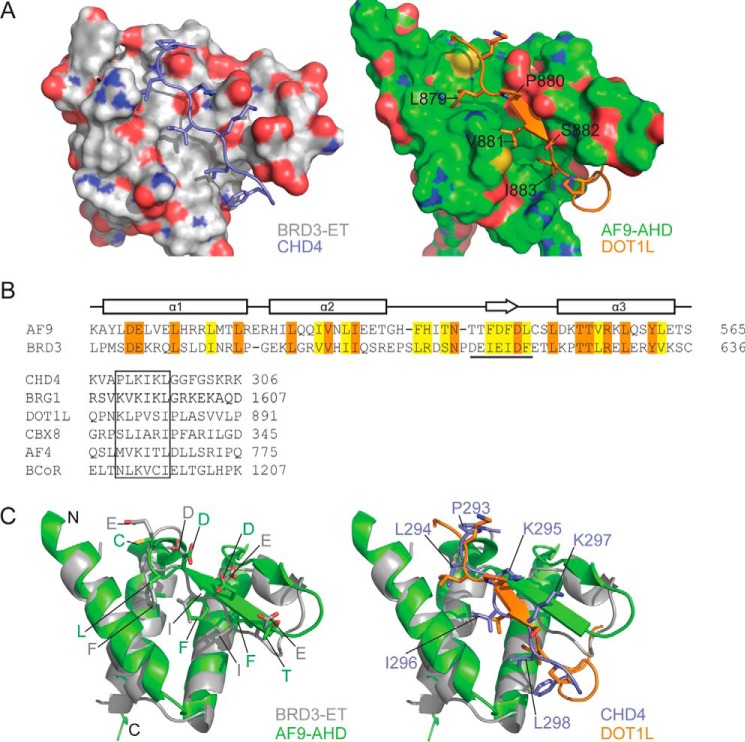
Members of the bromodomain and extra-terminal domain (BET) family of proteins (bromodomain-containing (BRD) 2, 3, 4, and T) are widely expressed and highly conserved regulators of gene expression in eukaryotes. These proteins have been intimately linked to human disease, and more than a dozen clinical trials are currently underway to test BET-protein inhibitors as modulators of cancer. However, although it is clear that these proteins use their bromodomains to bind both histones and transcription factors bearing acetylated lysine residues, the molecular mechanisms by which BET family proteins regulate gene expression are not well defined. In particular, the functions of the other domains such as the ET domain have been less extensively studied. Here, we examine the properties of the ET domain of BRD3 as a protein/protein interaction module. Using a combination of pulldown and biophysical assays, we demonstrate that BRD3 binds to a range of chromatin-remodeling complexes, including the NuRD, BAF, and INO80 complexes, via a short linear "KIKL" motif in one of the complex subunits. NMR-based structural analysis revealed that, surprisingly, this mode of interaction is shared by the AF9 and ENL transcriptional coregulators that contain an acetyl-lysine-binding YEATS domain and regulate transcriptional elongation. This observation establishes a functional commonality between these two families of cancer-related transcriptional regulators. In summary, our data provide insight into the mechanisms by which BET family proteins might link chromatin acetylation to transcriptional outcomes and uncover an unexpected functional similarity between BET and YEATS family proteins.
Keywords: AF9; BET family proteins; acetylation; bromodomain; extraterminal domain; nuclear magnetic resonance (NMR); protein structure; protein/protein interaction; transcriptional coactivator.
© 2018 by The American Society for Biochemistry and Molecular Biology, Inc.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
Similar articles
-
Comparative structure-function analysis of bromodomain and extraterminal motif (BET) proteins in a gene-complementation system.J Biol Chem. 2020 Feb 14;295(7):1898-1914. doi: 10.1074/jbc.RA119.010679. Epub 2019 Dec 2. J Biol Chem. 2020. PMID: 31792058 Free PMC article.
-
Multivalent nucleosome scaffolding by bromodomain and extraterminal domain tandem bromodomains.J Biol Chem. 2025 Mar;301(3):108289. doi: 10.1016/j.jbc.2025.108289. Epub 2025 Feb 10. J Biol Chem. 2025. PMID: 39938804 Free PMC article.
-
Selective recognition of acetylated histones by bromodomains in transcriptional co-activators.Biochem J. 2007 Feb 15;402(1):125-33. doi: 10.1042/BJ20060907. Biochem J. 2007. PMID: 17049045 Free PMC article.
-
Therapeutic targeting of BET bromodomain and other epigenetic acetylrecognition domain-containing factors.Curr Opin Genet Dev. 2024 Jun;86:102181. doi: 10.1016/j.gde.2024.102181. Epub 2024 Apr 2. Curr Opin Genet Dev. 2024. PMID: 38564841 Review.
-
The Bromodomain and Extra-Terminal Domain (BET) Family: Functional Anatomy of BET Paralogous Proteins.Int J Mol Sci. 2016 Nov 7;17(11):1849. doi: 10.3390/ijms17111849. Int J Mol Sci. 2016. PMID: 27827996 Free PMC article. Review.
Cited by
-
Functional coordination of BET family proteins underlies altered transcription associated with memory impairment in fragile X syndrome.Sci Adv. 2021 May 19;7(21):eabf7346. doi: 10.1126/sciadv.abf7346. Print 2021 May. Sci Adv. 2021. PMID: 34138732 Free PMC article.
-
Structure Determination of Challenging Protein-Peptide Complexes Combining NMR Chemical Shift Data and Molecular Dynamics Simulations.J Chem Inf Model. 2023 Apr 10;63(7):2058-2072. doi: 10.1021/acs.jcim.2c01595. Epub 2023 Mar 29. J Chem Inf Model. 2023. PMID: 36988562 Free PMC article.
-
Rerouting therapeutic peptides and unlocking their potential against SARS-CoV2.3 Biotech. 2025 May;15(5):116. doi: 10.1007/s13205-025-04270-0. Epub 2025 Apr 4. 3 Biotech. 2025. PMID: 40191455 Review.
-
Order in Disorder as Observed by the "Hydrophobic Cluster Analysis" of Protein Sequences.Proteomics. 2018 Nov;18(21-22):e1800054. doi: 10.1002/pmic.201800054. Epub 2018 Oct 30. Proteomics. 2018. PMID: 30299594 Free PMC article. Review.
-
Improving TRAIL-induced apoptosis in cancers by interfering with histone modifications.Cancer Drug Resist. 2020 Oct 9;3(4):791-803. doi: 10.20517/cdr.2020.58. eCollection 2020. Cancer Drug Resist. 2020. PMID: 35582230 Free PMC article. Review.
References
-
- Filippakopoulos P., Picaud S., Mangos M., Keates T., Lambert J.-P., Barsyte-Lovejoy D., Felletar I., Volkmer R., Müller S., Pawson T., Gingras A.-C., Arrowsmith C. H., and Knapp S. (2012) Histone recognition and large-scale structural analysis of the human bromodomain family. Cell 149, 214–231 10.1016/j.cell.2012.02.013 - DOI - PMC - PubMed
-
- Cheung K. L., Zhang F., Jaganathan A., Sharma R., Zhang Q., Konuma T., Shen T., Lee J.-Y., Ren C., Chen C.-H., Lu G., Olson M. R., Zhang W., Kaplan M. H., Littman D. R., et al. (2017) Distinct roles of Brd2 and Brd4 in potentiating the transcriptional program for Th17 cell differentiation. Mol. Cell 65, 1068–1080 10.1016/j.molcel.2016.12.022 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
