

Molecular Camouflage of Plasmodium falciparum Merozoites by Binding of Host Vitronectin to P47 Fragment of SERA5
- PMID: 29567995
- PMCID: PMC5864917
- DOI: 10.1038/s41598-018-23194-9
Molecular Camouflage of Plasmodium falciparum Merozoites by Binding of Host Vitronectin to P47 Fragment of SERA5
Abstract
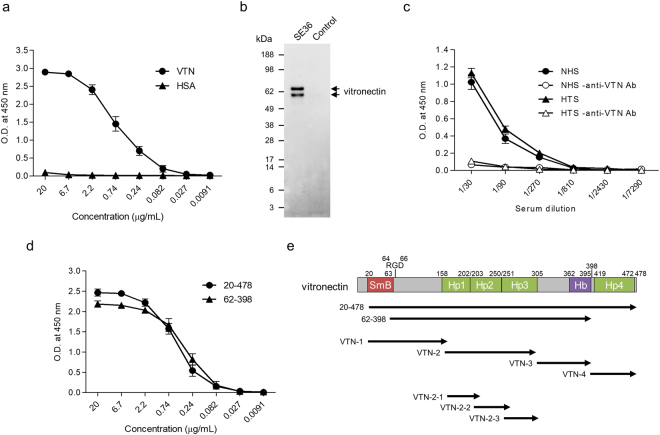
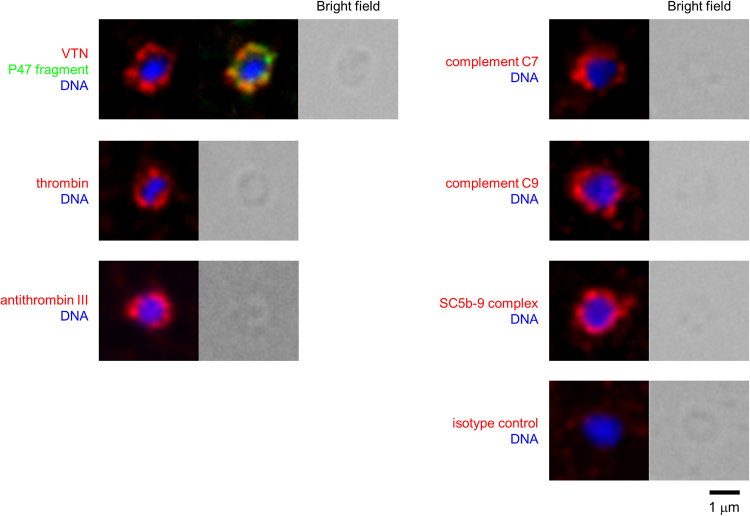
The malaria parasite Plasmodium falciparum proliferates in the blood stream where the host immune system is most active. To escape from host immunity, P. falciparum has developed a number of evasion mechanisms. Serine repeat antigen 5 (SERA5) is a blood stage antigen highly expressed at late trophozoite and schizont stages. The P47 N-terminal domain of SERA5, the basis of SE36 antigen of the blood stage vaccine candidate under clinical trials, covers the merozoite surface. Exploring the role of the P47 domain, screening of serum proteins showed that vitronectin (VTN) directly binds to 20 residues in the C-terminal region of SE36. VTN co-localized with P47 domain in the schizont and merozoite stages. Phagocytosis assay using THP-1 cells demonstrated that VTN bound to SE36 prevented engulfment of SE36-beads. In addition, several serum proteins localized on the merozoite surface, suggesting that host proteins camouflage merozoites against host immunity via binding to VTN.
Conflict of interest statement
The authors declare no competing interests.
Figures




Similar articles
-
Plasmodium falciparum MSP3 Exists in a Complex on the Merozoite Surface and Generates Antibody Response during Natural Infection.Infect Immun. 2018 Jul 23;86(8):e00067-18. doi: 10.1128/IAI.00067-18. Print 2018 Aug. Infect Immun. 2018. PMID: 29760216 Free PMC article.
-
Antibodies elicited by a virosomally formulated Plasmodium falciparum serine repeat antigen-5 derived peptide detect the processed 47 kDa fragment both in sporozoites and merozoites.Peptides. 2007 Oct;28(10):2051-60. doi: 10.1016/j.peptides.2007.08.007. Epub 2007 Aug 17. Peptides. 2007. PMID: 17875342
-
A novel Plasmodium falciparum erythrocyte binding protein associated with the merozoite surface, PfDBLMSP.Int J Parasitol. 2009 Jun;39(7):763-73. doi: 10.1016/j.ijpara.2008.12.004. Int J Parasitol. 2009. PMID: 19367830
-
Antigenic Variation in Plasmodium falciparum.Results Probl Cell Differ. 2015;57:47-90. doi: 10.1007/978-3-319-20819-0_3. Results Probl Cell Differ. 2015. PMID: 26537377 Review.
-
Plasmodium falciparum serine repeat antigen 5 (SE36) as a malaria vaccine candidate.Vaccine. 2011 Aug 11;29(35):5837-45. doi: 10.1016/j.vaccine.2011.06.052. Epub 2011 Jun 28. Vaccine. 2011. PMID: 21718740 Review.
Cited by
-
Plasmodium falciparum Blood Stage Antimalarial Vaccines: An Analysis of Ongoing Clinical Trials and New Perspectives Related to Synthetic Vaccines.Front Microbiol. 2019 Dec 3;10:2712. doi: 10.3389/fmicb.2019.02712. eCollection 2019. Front Microbiol. 2019. PMID: 31849871 Free PMC article. Review.
-
Holotomography: Refractive Index as an Intrinsic Imaging Contrast for 3-D Label-Free Live Cell Imaging.Adv Exp Med Biol. 2021;1310:211-238. doi: 10.1007/978-981-33-6064-8_10. Adv Exp Med Biol. 2021. PMID: 33834439
-
Microbial evasion of the complement system: a continuous and evolving story.Front Immunol. 2024 Jan 4;14:1281096. doi: 10.3389/fimmu.2023.1281096. eCollection 2023. Front Immunol. 2024. PMID: 38239357 Free PMC article. Review.
-
Three-dimensional label-free observation of individual bacteria upon antibiotic treatment using optical diffraction tomography.Biomed Opt Express. 2020 Feb 3;11(3):1257-1267. doi: 10.1364/BOE.377740. eCollection 2020 Mar 1. Biomed Opt Express. 2020. PMID: 32206407 Free PMC article.
-
Characteristic features of the SERA multigene family in the malaria parasite.Parasit Vectors. 2020 Apr 6;13(1):170. doi: 10.1186/s13071-020-04044-y. Parasit Vectors. 2020. PMID: 32252804 Free PMC article. Review.
References
-
- Pérez Arellano JL, Espinoza EY, Sánchez MM, Muro A. Evasion mechanisms of parasites. Revista Ibérica de Parasitología. 2001;61:4–16.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
