

Prior Expectation Modulates Repetition Suppression without Perceptual Awareness
- PMID: 29568041
- PMCID: PMC5864919
- DOI: 10.1038/s41598-018-23467-3
Prior Expectation Modulates Repetition Suppression without Perceptual Awareness
Abstract
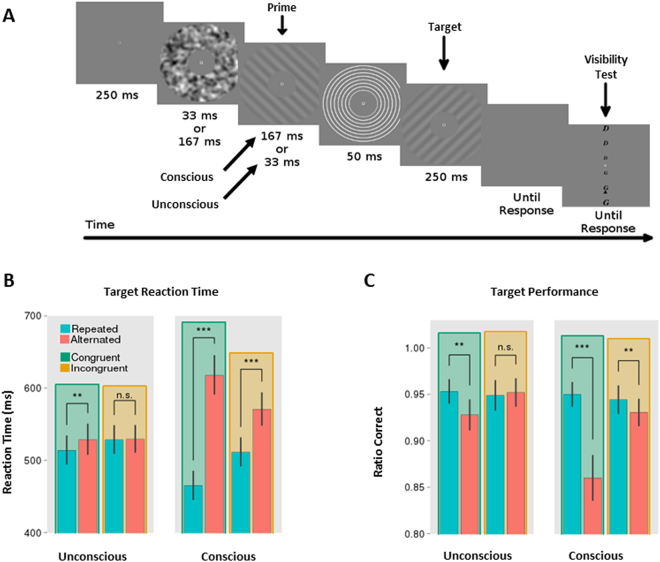
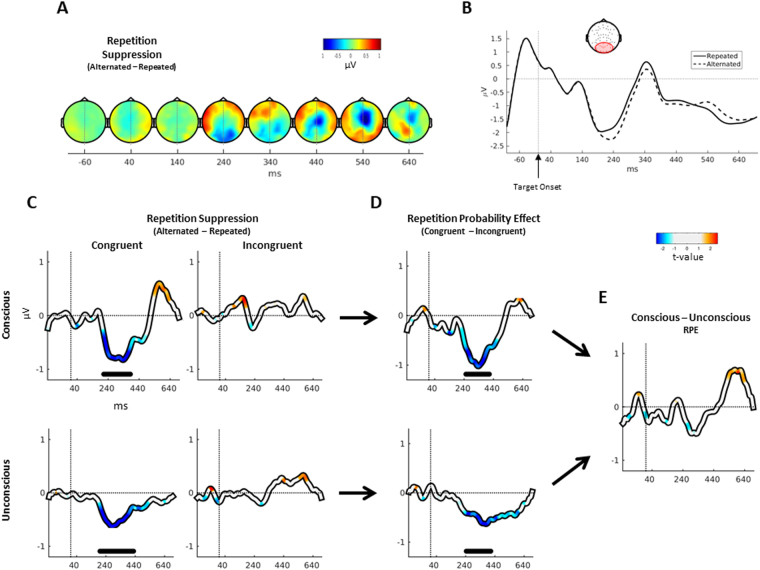
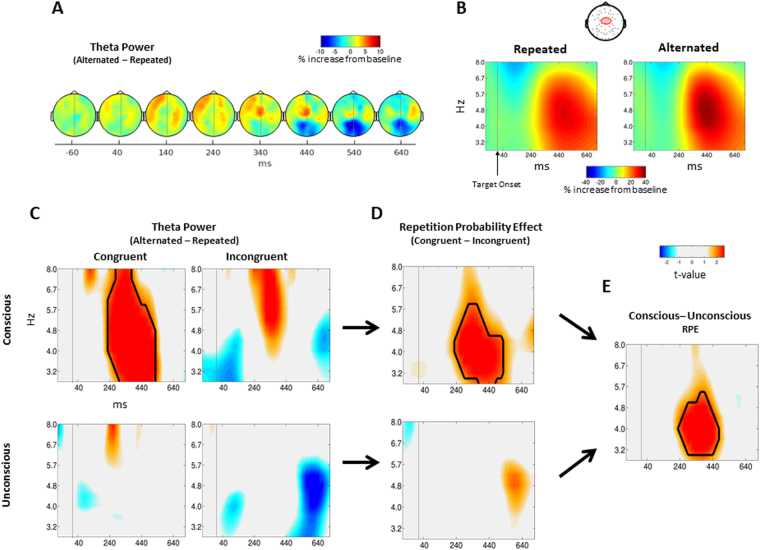
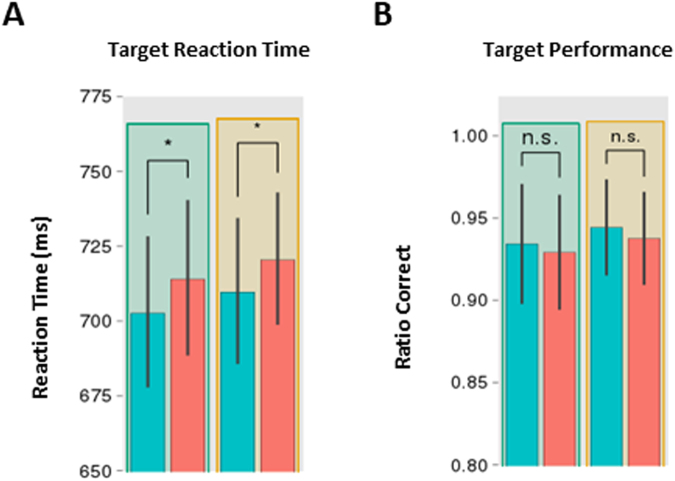
Stimulus repetition induces attenuated brain responses. This phenomenon, termed repetition suppression (RS), is classically held to stem from bottom-up neuronal adaptation. However, recent studies suggest that RS is driven by top-down predictive mechanisms. It remains controversial whether these top-down mechanisms of RS rely on conscious strategies, or if they represent a more fundamental aspect of perception, coding for physical properties of the repeated feature. The presence of top-down effects in the absence of perceptual awareness would indicate that conscious strategies are not sufficient to explain top-down mechanisms of RS. We combined an unconscious priming paradigm with EEG recordings and tested whether RS can be modulated by the probability of encountering a repetition, even in the absence of awareness. Our results show that both behavioural priming and RS near occipital areas are modulated by repetition probability, regardless of prime awareness. This contradicts previous findings that have argued that RS modulation is a by-product of conscious strategies. In contrast, we found that the increase in theta-band power following unrepeated trials - an index of conflict detection - is modulated only by expectations during conscious primes, implicating the use of conscious strategies. Together, our results suggest that the influence of predictions on RS can be either automatic in sensory brain regions or dependent on conscious strategies.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Kourtzi, Z. & Grill-Spector, K. fMRI adaptation: a tool for studying visual representations in the primate brain. Fitting Mind to World Adapt. After-Effects High-Level Vis. (Rhodes, G. Clifford, C. W. G. eds) 173–188 (2005).
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
