

The NMR contribution to protein-protein networking in Fe-S protein maturation
- PMID: 29569085
- PMCID: PMC6006191
- DOI: 10.1007/s00775-018-1552-x
The NMR contribution to protein-protein networking in Fe-S protein maturation
Erratum in
-
Correction to: The NMR contribution to protein-protein networking in Fe-S protein maturation.J Biol Inorg Chem. 2018 Jun;23(4):687. doi: 10.1007/s00775-018-1573-5. J Biol Inorg Chem. 2018. PMID: 29855713 Free PMC article.
Abstract
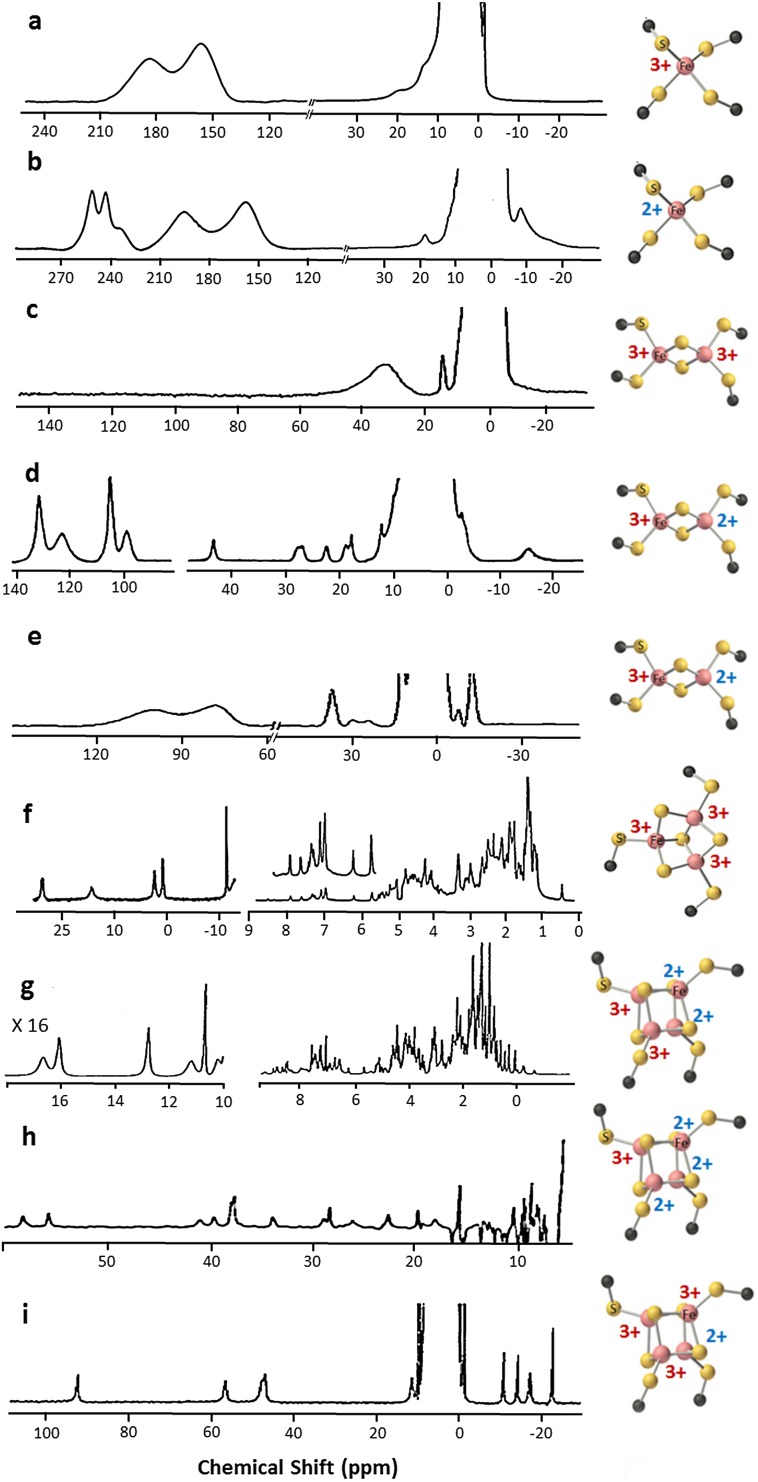
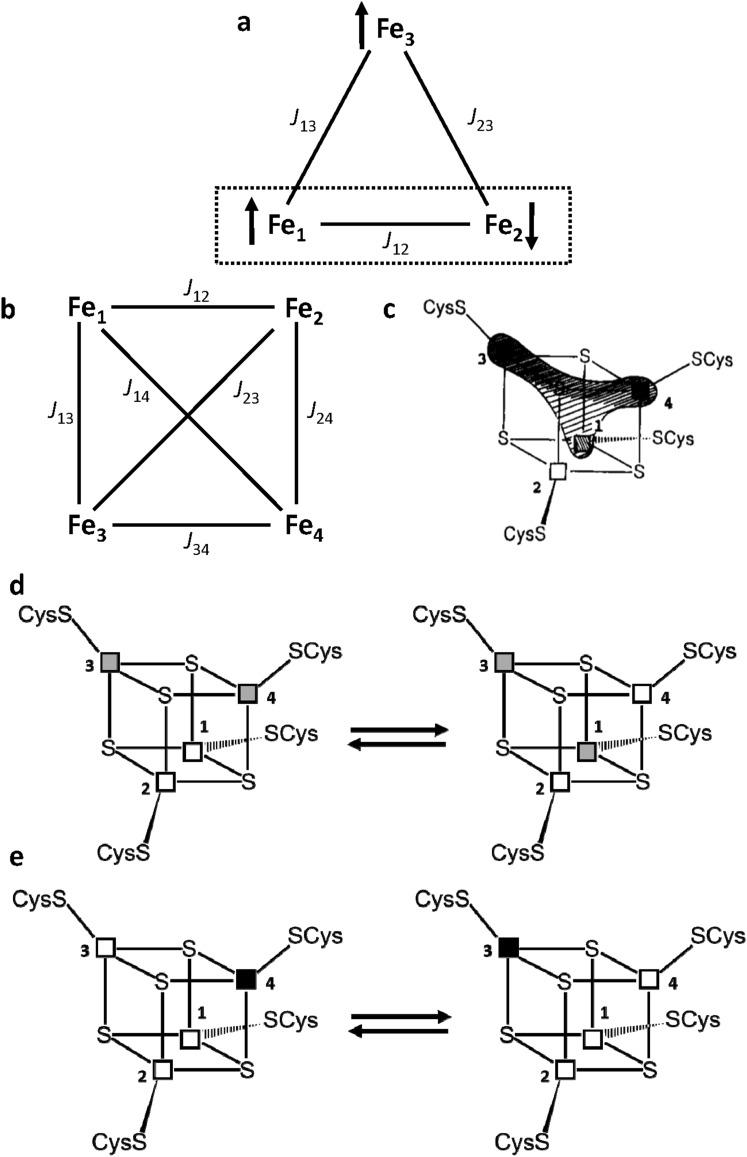
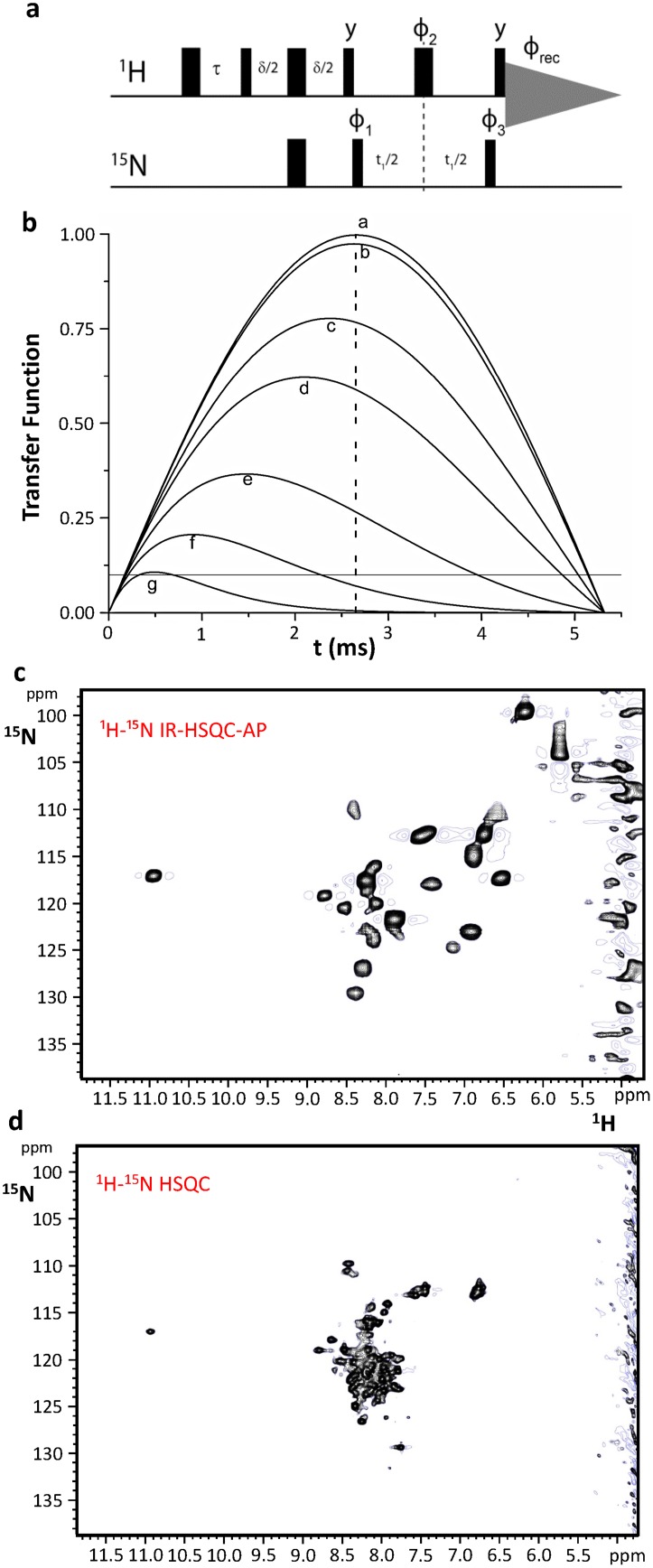
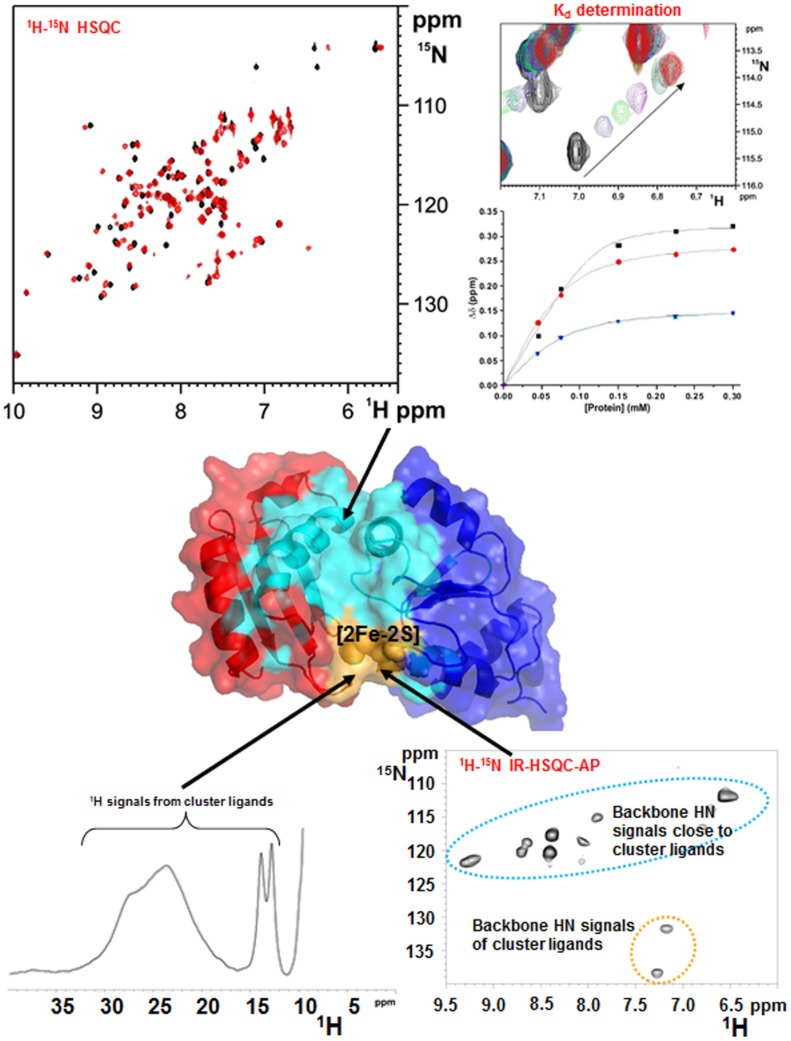
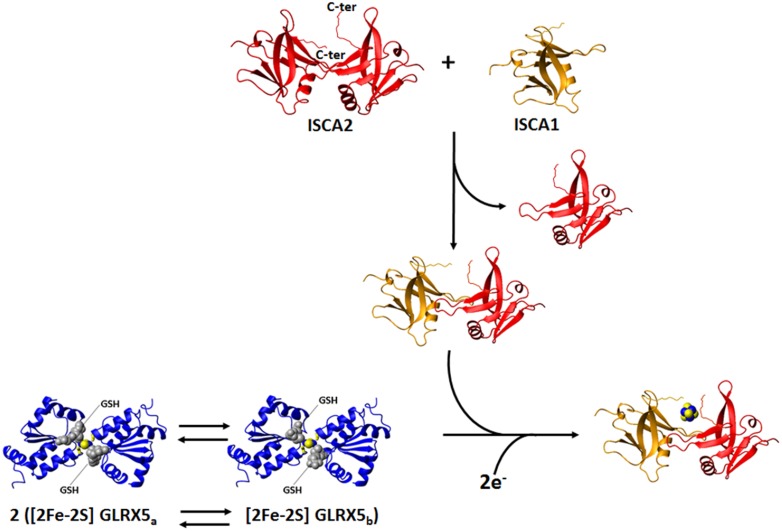
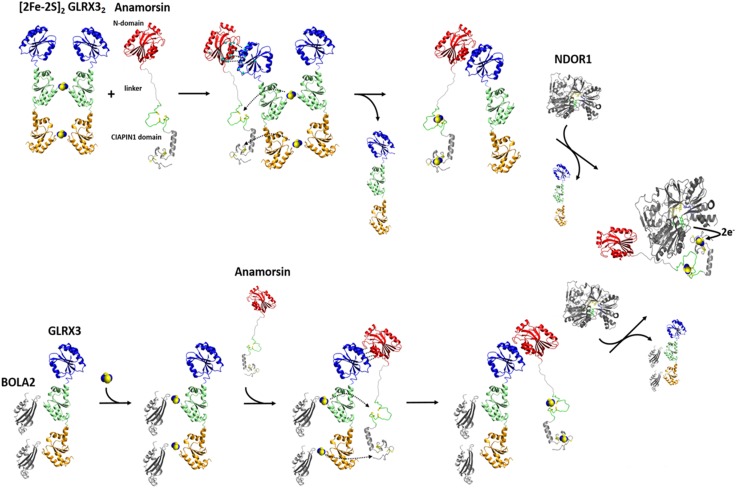
Iron-sulfur proteins were among the first class of metalloproteins that were actively studied using NMR spectroscopy tailored to paramagnetic systems. The hyperfine shifts, their temperature dependencies and the relaxation rates of nuclei of cluster-bound residues are an efficient fingerprint of the nature and the oxidation state of the Fe-S cluster. NMR significantly contributed to the analysis of the magnetic coupling patterns and to the understanding of the electronic structure occurring in [2Fe-2S], [3Fe-4S] and [4Fe-4S] clusters bound to proteins. After the first NMR structure of a paramagnetic protein was obtained for the reduced E. halophila HiPIP I, many NMR structures were determined for several Fe-S proteins in different oxidation states. It was found that differences in chemical shifts, in patterns of unobserved residues, in internal mobility and in thermodynamic stability are suitable data to map subtle changes between the two different oxidation states of the protein. Recently, the interaction networks responsible for maturing human mitochondrial and cytosolic Fe-S proteins have been largely characterized by combining solution NMR standard experiments with those tailored to paramagnetic systems. We show here the contribution of solution NMR in providing a detailed molecular view of "Fe-S interactomics". This contribution was particularly effective when protein-protein interactions are weak and transient, and thus difficult to be characterized at high resolution with other methodologies.
Keywords: CIA machinery; Fe–S proteins; Hyperfine interactions; ISC machinery; Interactomics; NMR spectroscopy.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
