

Transglutaminase 2: Friend or foe? The discordant role in neurons and astrocytes
- PMID: 29570839
- PMCID: PMC5980740
- DOI: 10.1002/jnr.24239
Transglutaminase 2: Friend or foe? The discordant role in neurons and astrocytes
Abstract
Members of the transglutaminase family catalyze the formation of isopeptide bonds between a polypeptide-bound glutamine and a low molecular weight amine (e.g., spermidine) or the ɛ-amino group of a polypeptide-bound lysine. Transglutaminase 2 (TG2), a prominent member of this family, is unique because in addition to being a transamidating enzyme, it exhibits numerous other activities. As a result, TG2 plays a role in many physiological processes, and its function is highly cell type specific and relies upon a number of factors, including conformation, cellular compartment location, and local concentrations of Ca2+ and guanine nucleotides. TG2 is the most abundant transglutaminase in the central nervous system (CNS) and plays a pivotal role in the CNS injury response. How TG2 affects the cell in response to an insult is strikingly different in astrocytes and neurons. In neurons, TG2 supports survival. Overexpression of TG2 in primary neurons protects against oxygen and glucose deprivation (OGD)-induced cell death and in vivo results in a reduction in infarct volume subsequent to a stroke. Knockdown of TG2 in primary neurons results in a loss of viability. In contrast, deletion of TG2 from astrocytes results in increased survival following OGD and improved ability to protect neurons from injury. Here, a brief overview of TG2 is provided, followed by a discussion of the role of TG2 in transcriptional regulation, cellular dynamics, and cell death. The differing roles TG2 plays in neurons and astrocytes are highlighted and compared to how TG2 functions in other cell types.
Keywords: astrocytes; cell death; cytoskeleton; injury; neurites; neurons; transcription regulation.
© 2018 Wiley Periodicals, Inc.
Conflict of interest statement
All the authors declare that they have no conflicts of interest
Figures
Similar articles
-
Depletion of astrocytic transglutaminase 2 improves injury outcomes.Mol Cell Neurosci. 2018 Oct;92:128-136. doi: 10.1016/j.mcn.2018.06.007. Epub 2018 Jun 30. Mol Cell Neurosci. 2018. PMID: 29969654 Free PMC article.
-
Complete transglutaminase 2 ablation results in reduced stroke volumes and astrocytes that exhibit increased survival in response to ischemia.Neurobiol Dis. 2012 Mar;45(3):1042-50. doi: 10.1016/j.nbd.2011.12.023. Epub 2011 Dec 14. Neurobiol Dis. 2012. PMID: 22198379 Free PMC article.
-
Transglutaminase 2 modulation of NF-κB signaling in astrocytes is independent of its ability to mediate astrocytic viability in ischemic injury.Brain Res. 2017 Aug 1;1668:1-11. doi: 10.1016/j.brainres.2017.05.009. Epub 2017 May 15. Brain Res. 2017. PMID: 28522262 Free PMC article.
-
Transglutaminase 2: a molecular Swiss army knife.Biochim Biophys Acta. 2012 Feb;1823(2):406-19. doi: 10.1016/j.bbamcr.2011.09.012. Epub 2011 Oct 10. Biochim Biophys Acta. 2012. PMID: 22015769 Free PMC article. Review.
-
Transglutaminase 2: an enigmatic enzyme with diverse functions.Trends Biochem Sci. 2002 Oct;27(10):534-9. doi: 10.1016/s0968-0004(02)02182-5. Trends Biochem Sci. 2002. PMID: 12368090 Review.
Cited by
-
Propofol Suppresses Microglia Inflammation by Targeting TGM2/NF-κB Signaling.J Immunol Res. 2021 Aug 24;2021:4754454. doi: 10.1155/2021/4754454. eCollection 2021. J Immunol Res. 2021. PMID: 34485533 Free PMC article.
-
Circulating anti-hypothalamus antibodies in celiac patients: tissue transglutaminase friend or foe?Immunol Res. 2023 Dec;71(6):839-848. doi: 10.1007/s12026-023-09394-0. Epub 2023 May 23. Immunol Res. 2023. PMID: 37221348 Free PMC article.
-
Role of Transglutaminase 2 in Cell Death, Survival, and Fibrosis.Cells. 2021 Jul 20;10(7):1842. doi: 10.3390/cells10071842. Cells. 2021. PMID: 34360011 Free PMC article. Review.
-
Deletion or Inhibition of Astrocytic Transglutaminase 2 Promotes Functional Recovery after Spinal Cord Injury.Cells. 2021 Oct 29;10(11):2942. doi: 10.3390/cells10112942. Cells. 2021. PMID: 34831164 Free PMC article.
-
Opening up about Tissue Transglutaminase: When Conformation Matters More than Enzymatic Activity.Med One. 2018;3(6):e180011. doi: 10.20900/mo.20180011. Epub 2018 Nov 22. Med One. 2018. PMID: 30662957 Free PMC article.
References
-
- Achyuthan KE, Greenberg CS. Identification of a guanosine triphosphate-binding site on guinea pig liver transglutaminase. Role of GTP and calcium ions in modulating activity. J Biol Chem. 1987;262(4):1901–1906. - PubMed
-
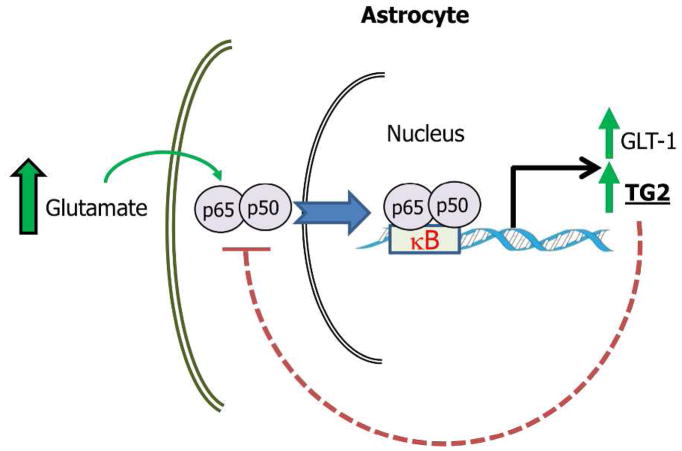
- Caccamo D, Campisi A, Curro M, Aguennouz M, Li Volti G, Avola R, Ientile R. Nuclear factor-kappab activation is associated with glutamate-evoked tissue transglutaminase up-regulation in primary astrocyte cultures. J Neurosci Res. 2005;82(6):858–865. - PubMed
-
- Campisi A, Caccamo D, Raciti G, Cannavo G, Macaione V, Curro M, Macaione S, Vanella A, Ientile R. Glutamate-induced increases in transglutaminase activity in primary cultures of astroglial cells. Brain research. 2003;978(1–2):24–30. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
