

Clock-dependent chromatin topology modulates circadian transcription and behavior
- PMID: 29572261
- PMCID: PMC5900709
- DOI: 10.1101/gad.312397.118
Clock-dependent chromatin topology modulates circadian transcription and behavior
Abstract
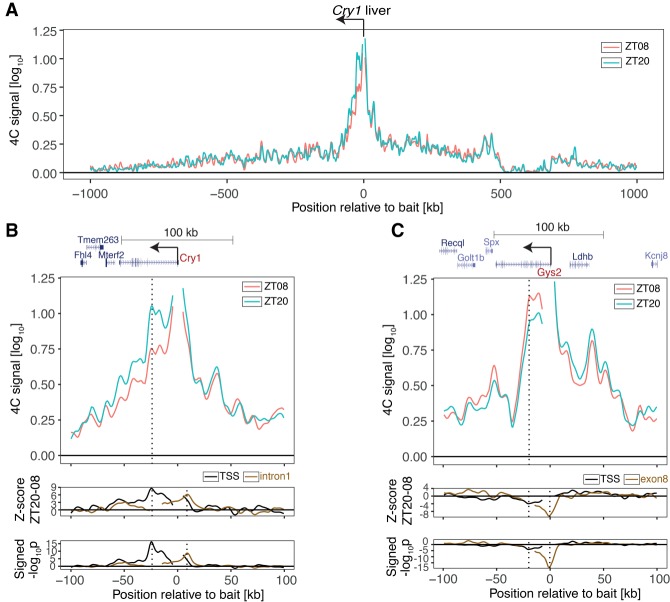
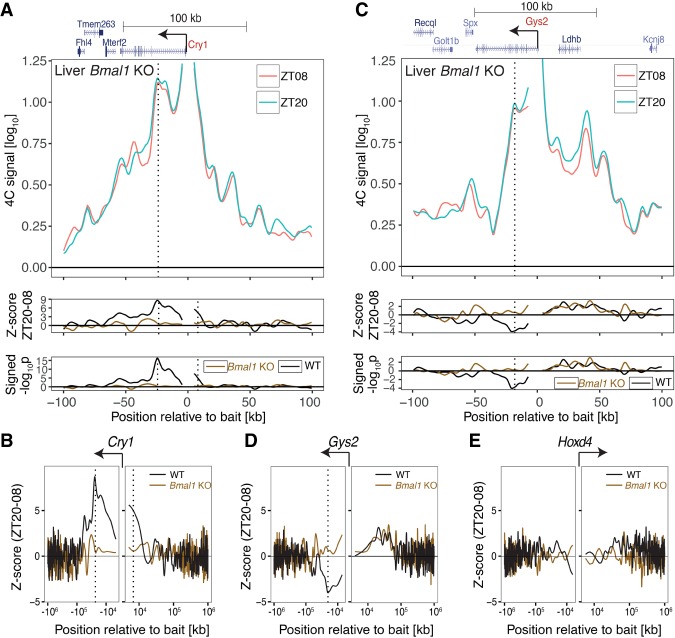
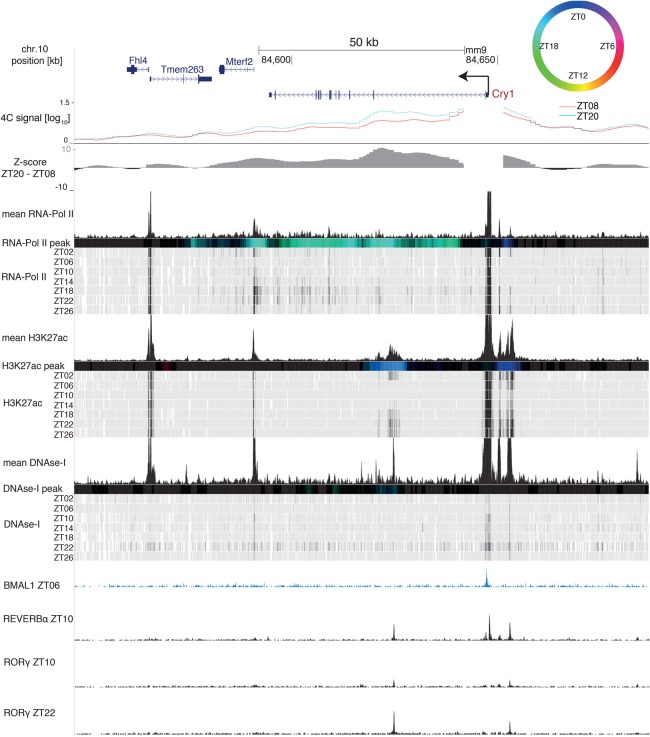
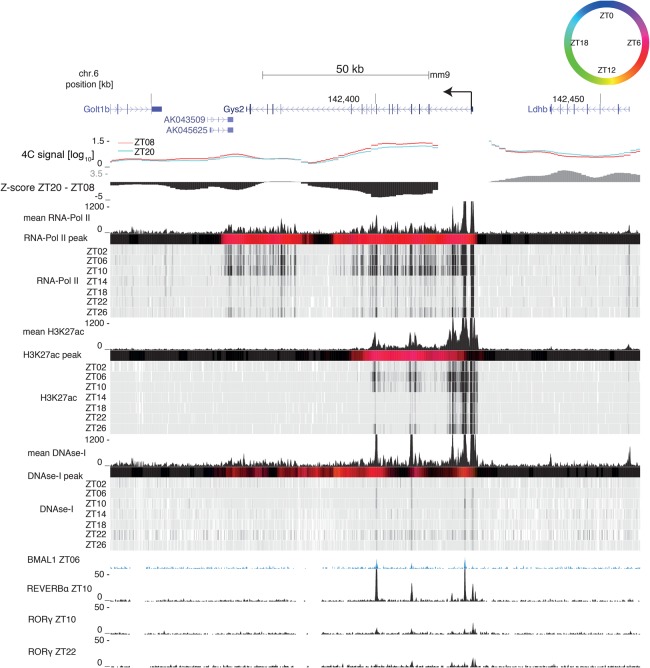
The circadian clock in animals orchestrates widespread oscillatory gene expression programs, which underlie 24-h rhythms in behavior and physiology. Several studies have shown the possible roles of transcription factors and chromatin marks in controlling cyclic gene expression. However, how daily active enhancers modulate rhythmic gene transcription in mammalian tissues is not known. Using circular chromosome conformation capture (4C) combined with sequencing (4C-seq), we discovered oscillatory promoter-enhancer interactions along the 24-h cycle in the mouse liver and kidney. Rhythms in chromatin interactions were abolished in arrhythmic Bmal1 knockout mice. Deleting a contacted intronic enhancer element in the Cryptochrome 1 (Cry1) gene was sufficient to compromise the rhythmic chromatin contacts in tissues. Moreover, the deletion reduced the daily dynamics of Cry1 transcriptional burst frequency and, remarkably, shortened the circadian period of locomotor activity rhythms. Our results establish oscillating and clock-controlled promoter-enhancer looping as a regulatory layer underlying circadian transcription and behavior.
Keywords: DNA regulatory elements; chromatin topology; circadian rhythms; promoter–enhancer loops; transcriptional bursting.
© 2018 Mermet et al.; Published by Cold Spring Harbor Laboratory Press.
Figures
References
-
- Allebrandt KV, Teder-Laving M, Akyol M, Pichler I, Muller-Myhsok B, Pramstaller P, Merrow M, Meitinger T, Metspalu A, Roenneberg T. 2010. CLOCK gene variants associate with sleep duration in two independent populations. Biol Psychiatry 67: 1040–1047. - PubMed
-
- Aschoff J, Pohl H. 1978. Phase relations between a circadian-rhythm and its Zeitgeber within range of entrainment. Naturwissenschaften 65: 80–84. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases