

Astrocytic glycogen metabolism in the healthy and diseased brain
- PMID: 29572349
- PMCID: PMC5950001
- DOI: 10.1074/jbc.R117.803239
Astrocytic glycogen metabolism in the healthy and diseased brain
Abstract
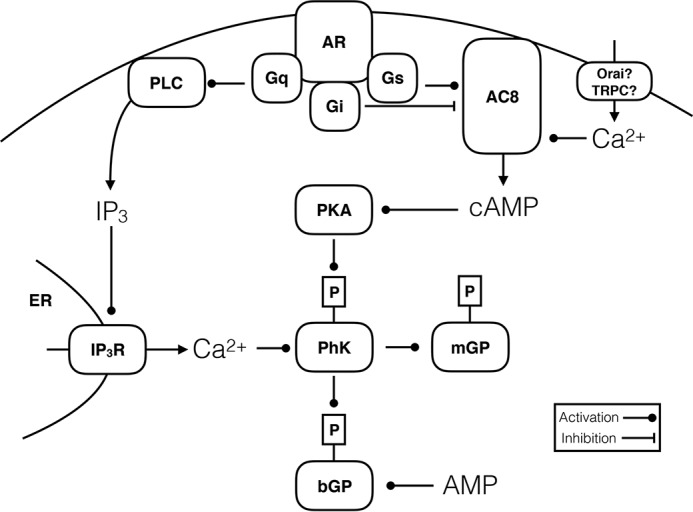
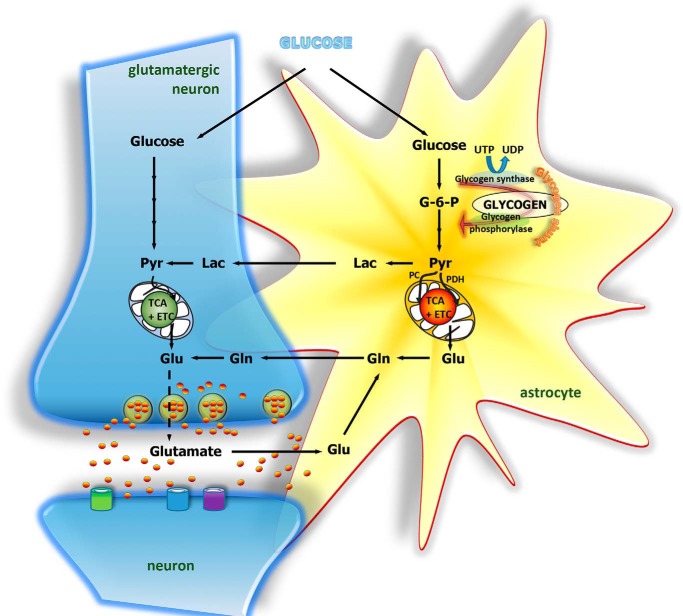
The brain contains a fairly low amount of glycogen, mostly located in astrocytes, a fact that has prompted the suggestion that glycogen does not have a significant physiological role in the brain. However, glycogen metabolism in astrocytes is essential for several key physiological processes and is adversely affected in disease. For instance, diminished ability to break down glycogen impinges on learning, and epilepsy, Alzheimer's disease, and type 2 diabetes are all associated with abnormal astrocyte glycogen metabolism. Glycogen metabolism supports astrocytic K+ and neurotransmitter glutamate uptake and subsequent glutamine synthesis-three fundamental steps in excitatory signaling at most brain synapses. Thus, there is abundant evidence for a key role of glycogen in brain function. Here, we summarize the physiological brain functions that depend on glycogen, discuss glycogen metabolism in disease, and investigate how glycogen breakdown is regulated at the cellular and molecular levels.
Keywords: astrocyte; brain; calcium; cyclic AMP (cAMP); disease; glycogen; neurological disease.
© 2018 by The American Society for Biochemistry and Molecular Biology, Inc.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
References
-
- Öz G., DiNuzzo M., Kumar A., Moheet A., Khowaja A., Kubisiak K., Eberly L. E., and Seaquist E. R. (2017) Cerebral glycogen in humans following acute and recurrent hypoglycemia: implications on a role in hypoglycemia unawareness. J. Cereb. Blood Flow Metab. 37, 2883–2893 10.1177/0271678X16678240 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
