

METHIONINE ADENOSYLTRANSFERASE4 Mediates DNA and Histone Methylation
- PMID: 29572390
- PMCID: PMC6001336
- DOI: 10.1104/pp.18.00183
METHIONINE ADENOSYLTRANSFERASE4 Mediates DNA and Histone Methylation
Abstract
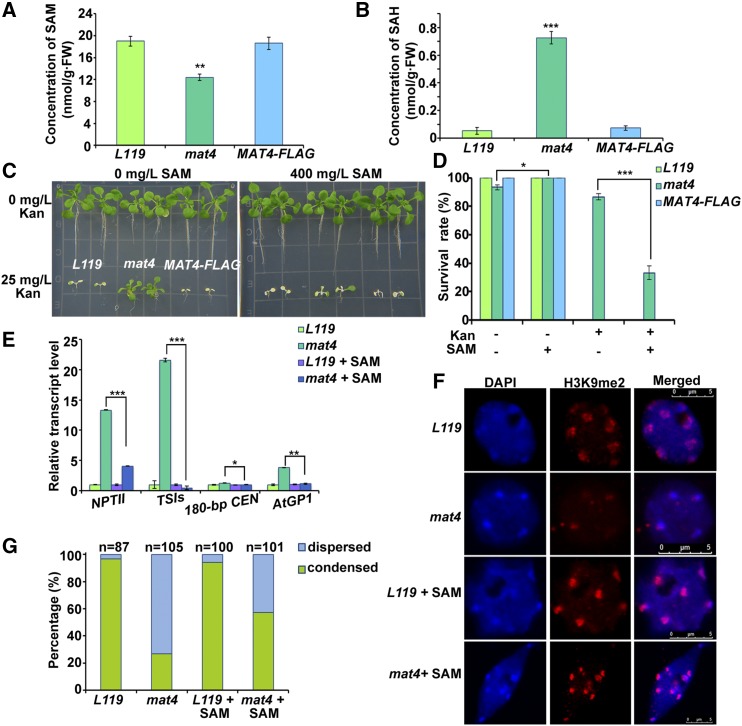
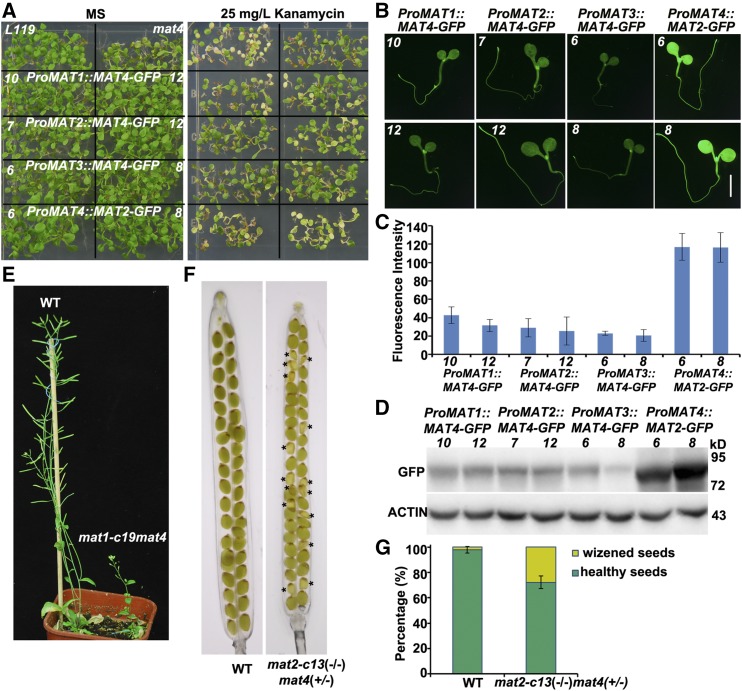
DNA and histone methylation coregulate heterochromatin formation and gene silencing in animals and plants. To identify factors involved in maintaining gene silencing, we conducted a forward genetic screen for mutants that release the silenced transgene Pro35S::NEOMYCIN PHOSPHOTRANSFERASE II in the transgenic Arabidopsis (Arabidopsis thaliana) line L119 We identified MAT4/SAMS3/MTO3/AT3G17390, which encodes methionine (Met) adenosyltransferase 4 (MAT4)/S-adenosyl-Met synthetase 3 that catalyzes the synthesis of S-adenosyl-Met (SAM) in the one-carbon metabolism cycle. mat4 mostly decreases CHG and CHH DNA methylation and histone H3K9me2 and reactivates certain silenced transposons. The exogenous addition of SAM partially rescues the epigenetic defects of mat4 SAM content and DNA methylation were reduced more in mat4 than in three other mat mutants. MAT4 knockout mutations generated by CRISPR/Cas9 were lethal, indicating that MAT4 is an essential gene in Arabidopsis. MAT1, 2, and 4 proteins exhibited nearly equal activity in an in vitro assay, whereas MAT3 exhibited higher activity. The native MAT4 promoter driving MAT1, 2, and 3 cDNA complemented the mat4 mutant. However, most mat4 transgenic lines carrying native MAT1, 2, and 3 promoters driving MAT4 cDNA did not complement the mat4 mutant because of their lower expression in seedlings. Genetic analyses indicated that the mat1mat4 double mutant is dwarfed and the mat2mat4 double mutant was nonviable, while mat1mat2 showed normal growth and fertility. These results indicate that MAT4 plays a predominant role in SAM production, plant growth, and development. Our findings provide direct evidence of the cooperative actions between metabolism and epigenetic regulation.
© 2018 American Society of Plant Biologists. All Rights Reserved.
Figures








Comment in
-
Powering Epigenetics through the 1C Pathway.Plant Physiol. 2018 Jun;177(2):443-444. doi: 10.1104/pp.18.00456. Plant Physiol. 2018. PMID: 29899054 Free PMC article. No abstract available.
References
-
- Cao X, Jacobsen SE (2002b) Role of the Arabidopsis DRM methyltransferases in de novo DNA methylation and gene silencing. Curr Biol 12: 1138–1144 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
