

A Proposed Circuit Computation in Basal Ganglia: History-Dependent Gain
- PMID: 29575303
- PMCID: PMC6001446
- DOI: 10.1002/mds.27321
A Proposed Circuit Computation in Basal Ganglia: History-Dependent Gain
Abstract
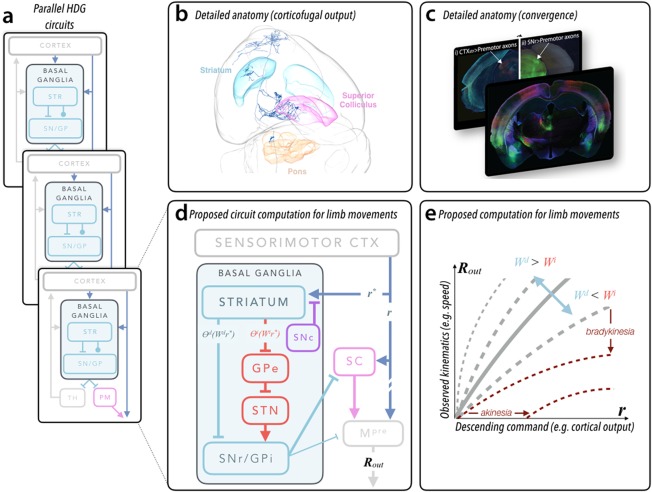
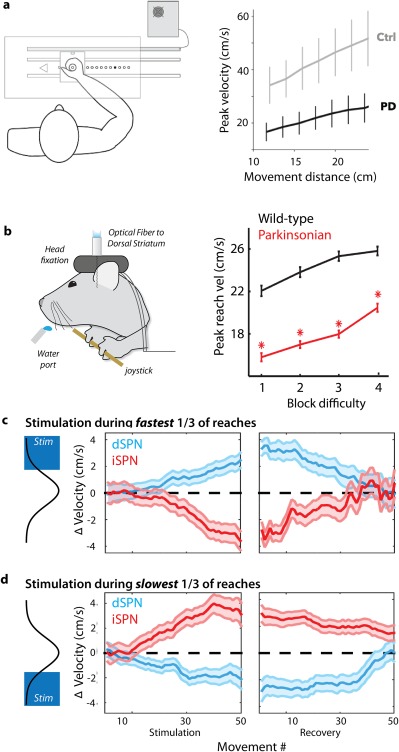
In this Scientific Perspectives we first review the recent advances in our understanding of the functional architecture of basal ganglia circuits. Then we argue that these data can best be explained by a model in which basal ganglia act to control the gain of movement kinematics to shape performance based on prior experience, which we refer to as a history-dependent gain computation. Finally, we discuss how insights from the history-dependent gain model might translate from the bench to the bedside, primarily the implications for the design of adaptive deep brain stimulation. Thus, we explicate the key empirical and conceptual support for a normative, computational model with substantial explanatory power for the broad role of basal ganglia circuits in health and disease. © 2018 The Authors. Movement Disorders published by Wiley Periodicals, Inc. on behalf of International Parkinson and Movement Disorder Society.
Keywords: basal ganglia; bradykinesia; dopamine; mouse models; neural circuits.
© 2018 The Authors. Movement Disorders published by Wiley Periodicals, Inc. on behalf of International Parkinson and Movement Disorder Society.
Figures
Similar articles
-
A possible correlation between the basal ganglia motor function and the inverse kinematics calculation.J Comput Neurosci. 2017 Dec;43(3):295-318. doi: 10.1007/s10827-017-0665-5. Epub 2017 Oct 23. J Comput Neurosci. 2017. PMID: 29063491
-
The Basal Ganglia in Action.Neuroscientist. 2017 Jun;23(3):299-313. doi: 10.1177/1073858416654115. Epub 2016 Jun 15. Neuroscientist. 2017. PMID: 27306757 Free PMC article. Review.
-
Modeling the functional organization of the basal ganglia. A parallel distributed processing approach.Mov Disord. 1991;6(3):189-204. doi: 10.1002/mds.870060302. Mov Disord. 1991. PMID: 1922123 Review.
-
The many worlds hypothesis of dopamine prediction error: implications of a parallel circuit architecture in the basal ganglia.Curr Opin Neurobiol. 2017 Oct;46:241-247. doi: 10.1016/j.conb.2017.08.015. Epub 2017 Oct 3. Curr Opin Neurobiol. 2017. PMID: 28985550 Review.
-
Functional Anatomy of Basal Ganglia Circuits with the Cerebral Cortex and the Cerebellum.Prog Neurol Surg. 2018;33:50-61. doi: 10.1159/000480748. Epub 2018 Jan 12. Prog Neurol Surg. 2018. PMID: 29332073 Review.
Cited by
-
Repeating patterns: Predictive processing suggests an aesthetic learning role of the basal ganglia in repetitive stereotyped behaviors.Front Psychol. 2022 Sep 8;13:930293. doi: 10.3389/fpsyg.2022.930293. eCollection 2022. Front Psychol. 2022. PMID: 36160532 Free PMC article.
-
Current approaches to characterize micro- and macroscale circuit mechanisms of Parkinson's disease in rodent models.Exp Neurol. 2022 May;351:114008. doi: 10.1016/j.expneurol.2022.114008. Epub 2022 Feb 9. Exp Neurol. 2022. PMID: 35149118 Free PMC article. Review.
-
Correlates of head-fixed orienting movements in mouse superior colliculus and substantia nigra pars reticulata.bioRxiv [Preprint]. 2025 May 8:2025.05.02.651955. doi: 10.1101/2025.05.02.651955. bioRxiv. 2025. PMID: 40654913 Free PMC article. Preprint.
-
Anterior Cingulate Cortex Contributes to the Hyperlocomotion under Nitrogen Narcosis.Neurosci Bull. 2025 May;41(5):775-789. doi: 10.1007/s12264-024-01278-z. Epub 2024 Aug 19. Neurosci Bull. 2025. PMID: 39158823
-
Distinct Populations of Motor Thalamic Neurons Encode Action Initiation, Action Selection, and Movement Vigor.J Neurosci. 2018 Jul 18;38(29):6563-6573. doi: 10.1523/JNEUROSCI.0463-18.2018. Epub 2018 Jun 22. J Neurosci. 2018. PMID: 29934350 Free PMC article.
References
-
- Meyer A, Hierons R. A Note on Thomas Willis' Views on the Corpus Striatum and the Internal Capsule. J Neurol Sci 1964;1(6):547‐554. - PubMed
-
- Jankovic J. Parkinson's disease: clinical features and diagnosis. J Neurol Neurosurg Psychiatry 2008;79(4):368‐376. - PubMed
-
- Kish SJ, Shannak K, Hornykiewicz O. Uneven pattern of dopamine loss in the striatum of patients with idiopathic Parkinson's disease. Pathophysiologic and clinical implications. N Engl J Med 1988;318(14):876‐880. - PubMed
-
- Dudman JT, Gerfen CR, The Basal Ganglia, in the Rat Nervous System. Paxinos G, ed. Amsterdam: Elsevier; 2015.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
