

Population and evolutionary dynamics in spatially structured seasonally varying environments
- PMID: 29575449
- PMCID: PMC6849584
- DOI: 10.1111/brv.12409
Population and evolutionary dynamics in spatially structured seasonally varying environments
Abstract
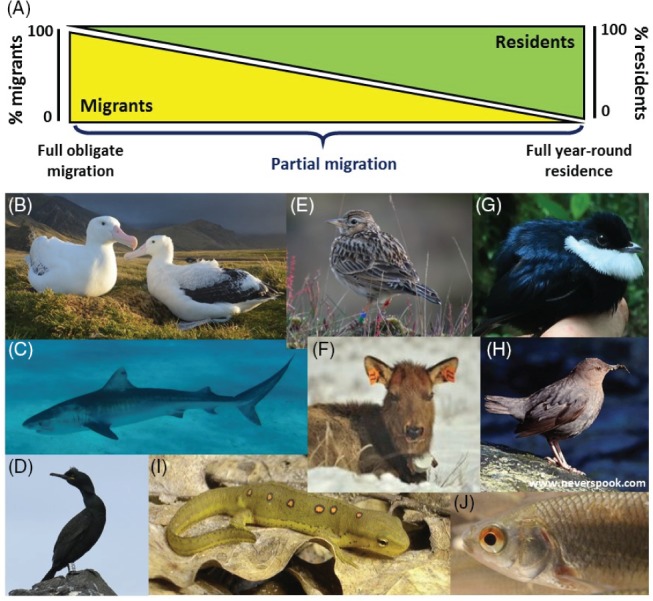
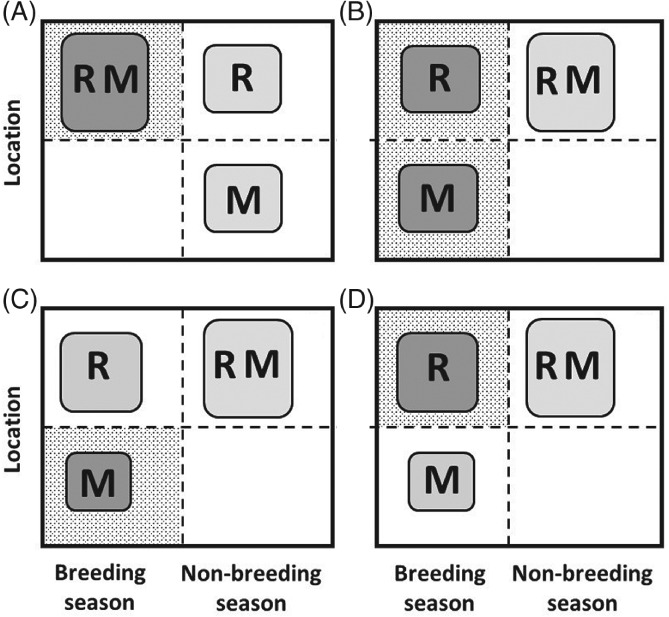
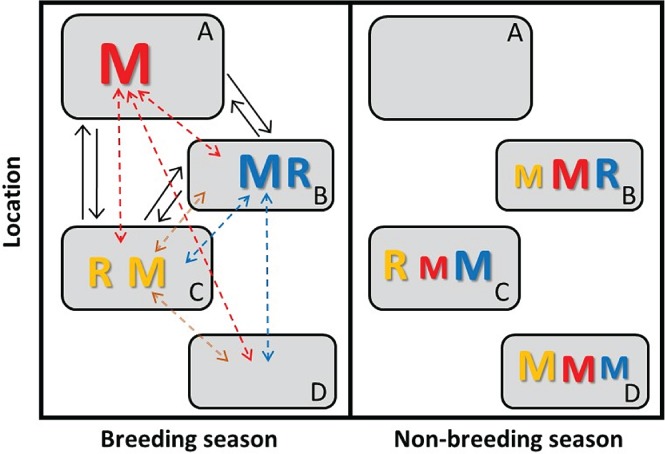
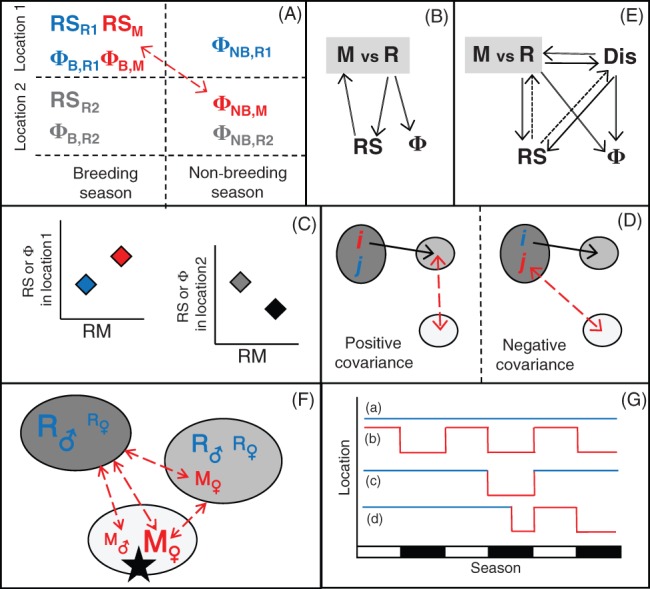
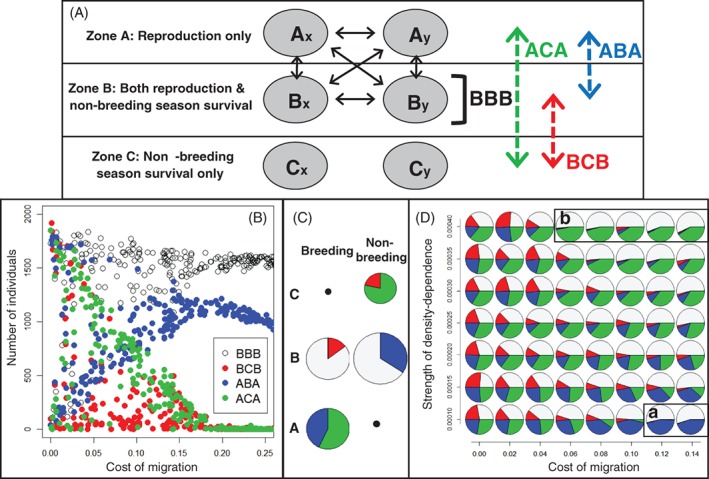
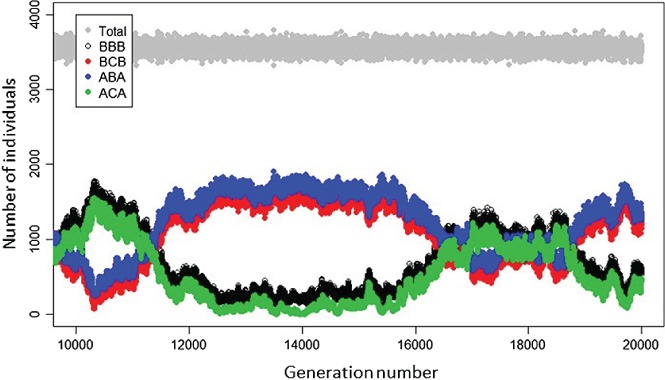
Increasingly imperative objectives in ecology are to understand and forecast population dynamic and evolutionary responses to seasonal environmental variation and change. Such population and evolutionary dynamics result from immediate and lagged responses of all key life-history traits, and resulting demographic rates that affect population growth rate, to seasonal environmental conditions and population density. However, existing population dynamic and eco-evolutionary theory and models have not yet fully encompassed within-individual and among-individual variation, covariation, structure and heterogeneity, and ongoing evolution, in a critical life-history trait that allows individuals to respond to seasonal environmental conditions: seasonal migration. Meanwhile, empirical studies aided by new animal-tracking technologies are increasingly demonstrating substantial within-population variation in the occurrence and form of migration versus year-round residence, generating diverse forms of 'partial migration' spanning diverse species, habitats and spatial scales. Such partially migratory systems form a continuum between the extreme scenarios of full migration and full year-round residence, and are commonplace in nature. Here, we first review basic scenarios of partial migration and associated models designed to identify conditions that facilitate the maintenance of migratory polymorphism. We highlight that such models have been fundamental to the development of partial migration theory, but are spatially and demographically simplistic compared to the rich bodies of population dynamic theory and models that consider spatially structured populations with dispersal but no migration, or consider populations experiencing strong seasonality and full obligate migration. Second, to provide an overarching conceptual framework for spatio-temporal population dynamics, we define a 'partially migratory meta-population' system as a spatially structured set of locations that can be occupied by different sets of resident and migrant individuals in different seasons, and where locations that can support reproduction can also be linked by dispersal. We outline key forms of within-individual and among-individual variation and structure in migration that could arise within such systems and interact with variation in individual survival, reproduction and dispersal to create complex population dynamics and evolutionary responses across locations, seasons, years and generations. Third, we review approaches by which population dynamic and eco-evolutionary models could be developed to test hypotheses regarding the dynamics and persistence of partially migratory meta-populations given diverse forms of seasonal environmental variation and change, and to forecast system-specific dynamics. To demonstrate one such approach, we use an evolutionary individual-based model to illustrate that multiple forms of partial migration can readily co-exist in a simple spatially structured landscape. Finally, we summarise recent empirical studies that demonstrate key components of demographic structure in partial migration, and demonstrate diverse associations with reproduction and survival. We thereby identify key theoretical and empirical knowledge gaps that remain, and consider multiple complementary approaches by which these gaps can be filled in order to elucidate population dynamic and eco-evolutionary responses to spatio-temporal seasonal environmental variation and change.
Keywords: demographic structure; density-dependence; eco-evolutionary dynamics; life-history variation; meta-population; movement ecology; partial migration; plasticity; population viability; seasonal migrant; vital rate.
© 2018 The Authors. Biological Reviews published by John Wiley & Sons Ltd on behalf of Cambridge Philosophical Society.
Figures
Similar articles
-
Strong survival selection on seasonal migration versus residence induced by extreme climatic events.J Anim Ecol. 2021 Apr;90(4):796-808. doi: 10.1111/1365-2656.13410. Epub 2021 Jan 26. J Anim Ecol. 2021. PMID: 33340099
-
Modelling the responses of partially migratory metapopulations to changing seasonal migration rates: From theory to data.J Anim Ecol. 2022 Sep;91(9):1781-1796. doi: 10.1111/1365-2656.13748. Epub 2022 Jul 17. J Anim Ecol. 2022. PMID: 35633181 Free PMC article.
-
Among-individual and within-individual variation in seasonal migration covaries with subsequent reproductive success in a partially migratory bird.Proc Biol Sci. 2020 Jul 29;287(1931):20200928. doi: 10.1098/rspb.2020.0928. Epub 2020 Jul 22. Proc Biol Sci. 2020. PMID: 32693718 Free PMC article.
-
Multi-generational fitness legacies of natural immigration: theoretical and empirical perspectives and opportunities.Biol Rev Camb Philos Soc. 2025 Jun;100(3):1250-1271. doi: 10.1111/brv.13183. Epub 2025 Feb 17. Biol Rev Camb Philos Soc. 2025. PMID: 39957338 Review.
-
A long winter for the Red Queen: rethinking the evolution of seasonal migration.Biol Rev Camb Philos Soc. 2019 Jun;94(3):737-752. doi: 10.1111/brv.12476. Epub 2018 Nov 4. Biol Rev Camb Philos Soc. 2019. PMID: 30393938 Review.
Cited by
-
Quantifying phenology and migratory behaviours of hummingbirds using single-site dynamics and mark-detection analyses.Proc Biol Sci. 2022 Sep 14;289(1982):20220991. doi: 10.1098/rspb.2022.0991. Epub 2022 Sep 14. Proc Biol Sci. 2022. PMID: 36100019 Free PMC article.
-
The physiology of movement.Mov Ecol. 2020 Feb 4;8:5. doi: 10.1186/s40462-020-0192-2. eCollection 2020. Mov Ecol. 2020. PMID: 32042434 Free PMC article. Review.
-
Temperature and microclimate refugia use influence migratory timings of a threatened grassland bird.Mov Ecol. 2023 Dec 1;11(1):75. doi: 10.1186/s40462-023-00437-7. Mov Ecol. 2023. PMID: 38041190 Free PMC article.
-
Maternally transferred thyroid hormones and life-history variation in birds.J Anim Ecol. 2022 Jul;91(7):1489-1506. doi: 10.1111/1365-2656.13708. Epub 2022 May 7. J Anim Ecol. 2022. PMID: 35470435 Free PMC article.
-
Comparing two measures of phenological synchrony in a predator-prey interaction: Simpler works better.J Anim Ecol. 2020 Mar;89(3):745-756. doi: 10.1111/1365-2656.13143. Epub 2019 Dec 17. J Anim Ecol. 2020. PMID: 31691954 Free PMC article.
References
-
- Aben, J. , Bocedi, G. , Palmer, S. C. F. , Pellikka, P. , Strubbe, D. , Hallmann, C. , Travis, J. M. J. , Lens, L. & Matthysen, E. (2016). The importance of realistic dispersal models in conservation planning: application of a novel modelling platform to evaluate management scenarios in an Afrotropical biodiversity hotspot. Journal of Applied Ecology 53, 1055–1065. - PMC - PubMed
-
- Ådahl, E. , Lundberg, P. & Jonzén, N. (2006). From climate change to population change: the need to consider annual life cycles. Global Change Biology 12, 1627–1633.
-
- Anderson, A. M. , Novak, S. J. , Smith, J. F. , Steenhof, K. & Heath, J. A. (2015). Nesting phenology, mate choice, and genetic divergence within a partially migratory population of American kestrels. Auk 133, 99–109.
-
- Avgar, T. , Street, G. & Fryxell, J. M. (2014). On the adaptive benefits of mammal migration. Canadian Journal of Zoology 92, 481–490.
-
- Bailey, L. D. & Van de pol, M. (2016). Tackling extremes: challenges for ecological and evolutionary research on extreme climatic events. Journal of Animal Ecology 85, 85–96. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
