

The bHLH transcription factor GmPIB1 facilitates resistance to Phytophthora sojae in Glycine max
- PMID: 29579245
- PMCID: PMC5920285
- DOI: 10.1093/jxb/ery103
The bHLH transcription factor GmPIB1 facilitates resistance to Phytophthora sojae in Glycine max
Abstract
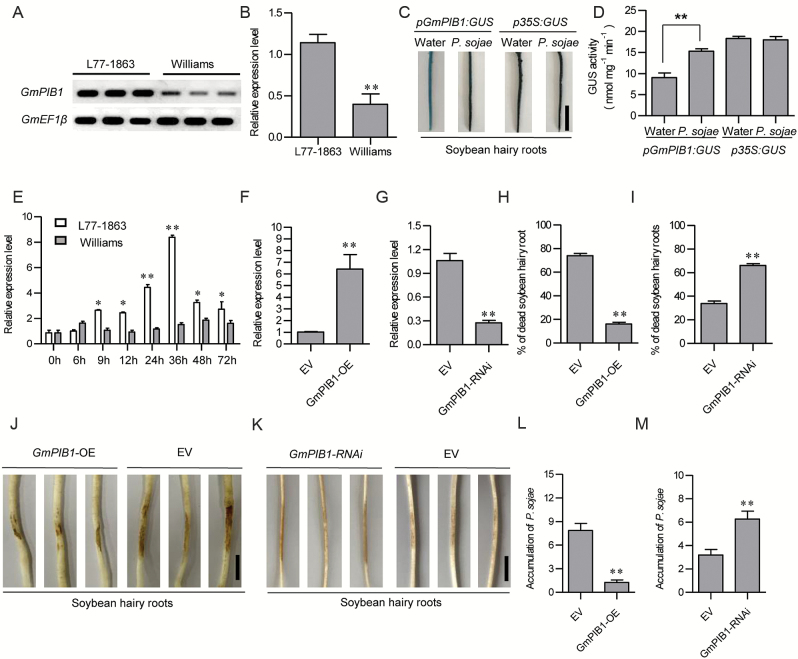
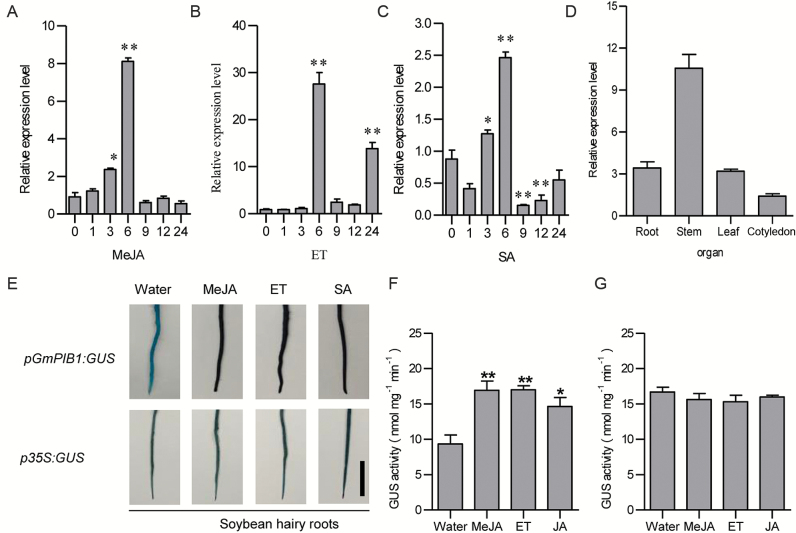
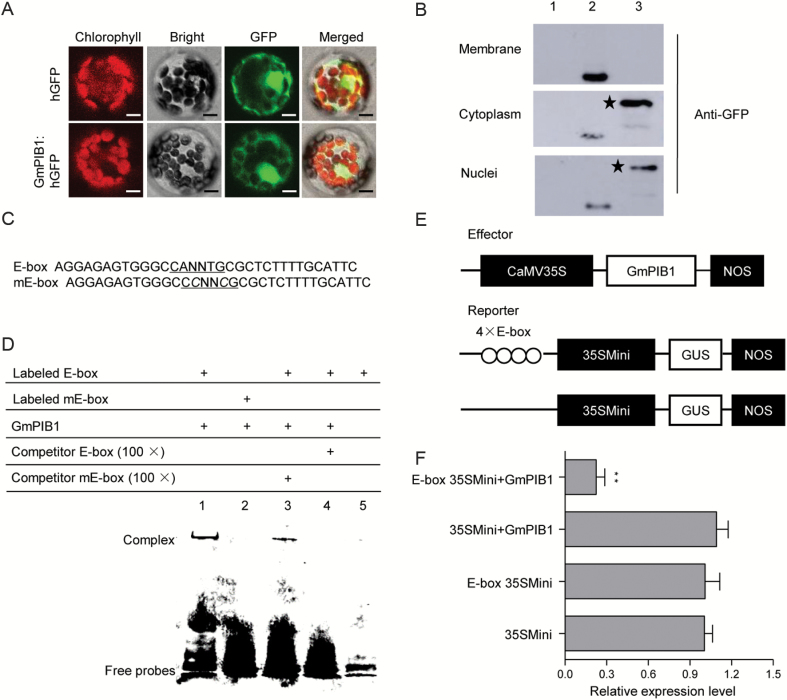
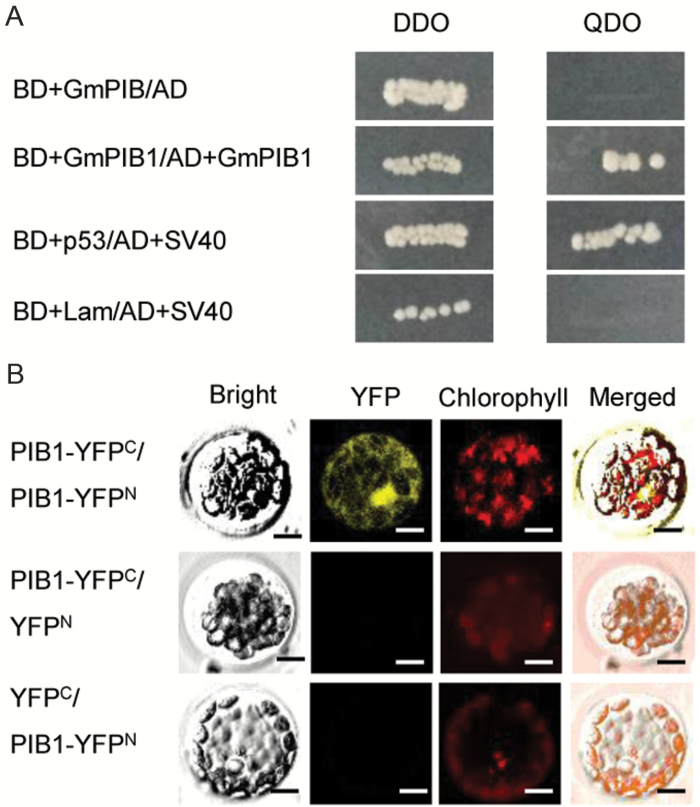
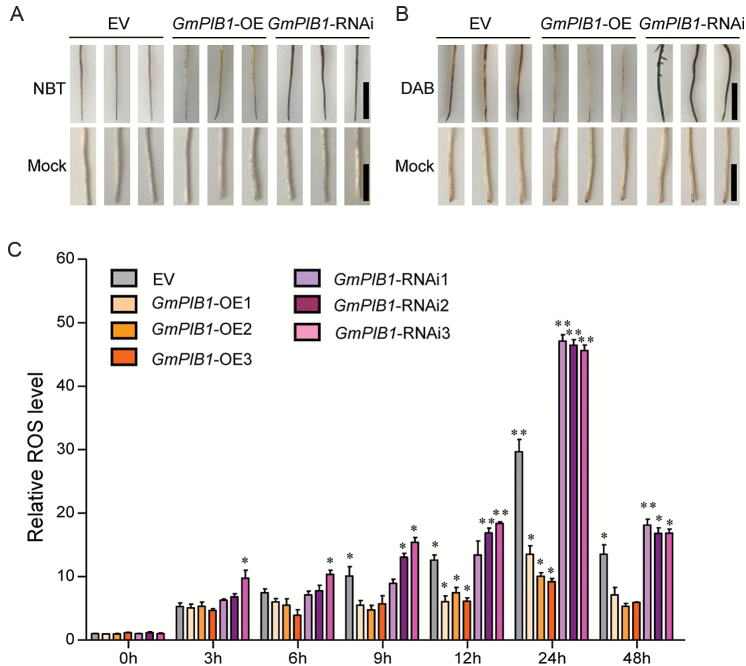
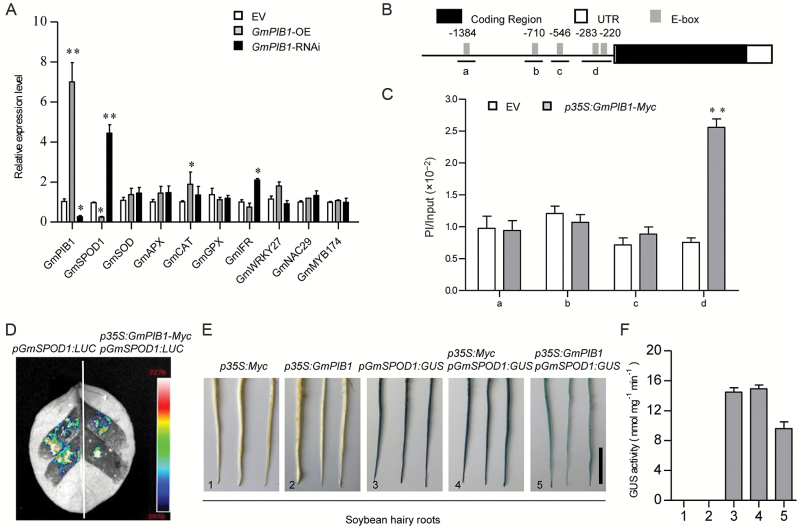
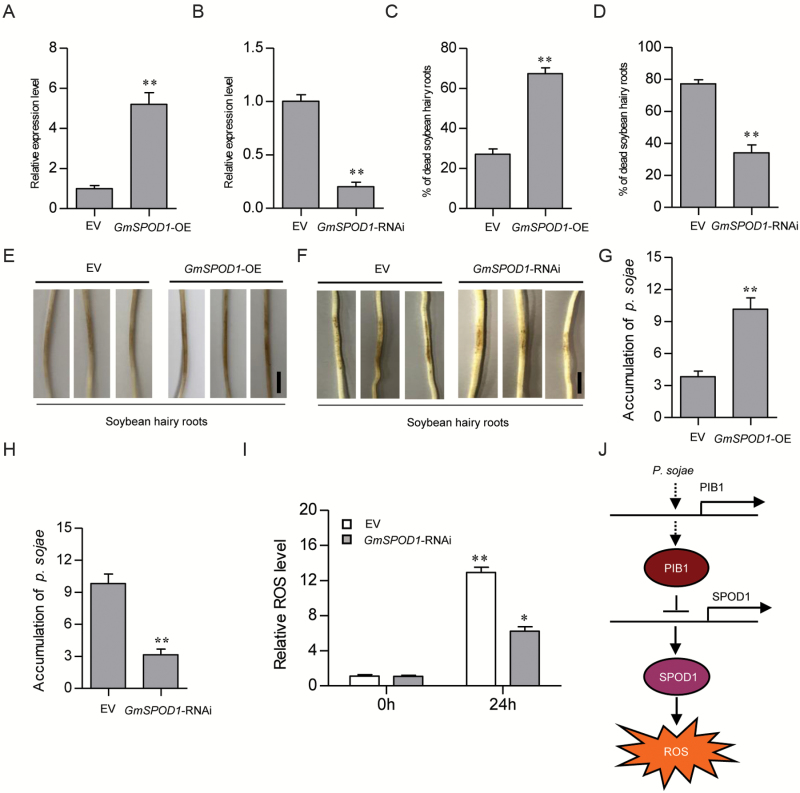
Phytophthora sojae Kaufmann and Gerdemann causes Phytophthora root rot, a destructive soybean disease worldwide. A basic helix-loop-helix (bHLH) transcription factor is thought to be involved in the response to P. sojae infection in soybean, as revealed by RNA sequencing (RNA-seq). However, the molecular mechanism underlying this response is currently unclear. Here, we explored the function and underlying mechanisms of a bHLH transcription factor in soybean, designated GmPIB1 (P. sojae-inducible bHLH transcription factor), during host responses to P. sojae. GmPIB1 was significantly induced by P. sojae in the resistant soybean cultivar 'L77-1863'. Analysis of transgenic soybean hairy roots with elevated or reduced expression of GmPIB1 demonstrated that GmPIB1 enhances resistance to P. sojae and reduces reactive oxygen species (ROS) accumulation. Quantitative reverse transcription PCR and chromatin immunoprecipitation-quantitative PCR assays revealed that GmPIB1 binds directly to the promoter of GmSPOD1 and represses its expression; this gene encodes a key enzyme in ROS production. Moreover, transgenic soybean hairy roots with GmSPOD1 silencing through RNA interference exhibited improved resistance to P. sojae and reduced ROS generation. These findings suggest that GmPIB1 enhances resistance to P. sojae by repressing the expression of GmSPOD1.
Figures
References
-
- Berrocal-Lobo M, Molina A, Solano R. 2002. Constitutive expression of ETHYLENE-RESPONSE-FACTOR1 in Arabidopsis confers resistance to several necrotrophic fungi. The Plant Journal 29, 23–32. - PubMed
-
- Blair JE, Coffey MD, Park SY, Geiser DM, Kang S. 2008. A multi-locus phylogeny for Phytophthora utilizing markers derived from complete genome sequences. Fungal Genetics and Biology 45, 266–277. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
