

Dose-Dependent Prevention of Metabolic and Neurologic Disease in Murine MPS II by ZFN-Mediated In Vivo Genome Editing
- PMID: 29580682
- PMCID: PMC6080131
- DOI: 10.1016/j.ymthe.2018.03.002
Dose-Dependent Prevention of Metabolic and Neurologic Disease in Murine MPS II by ZFN-Mediated In Vivo Genome Editing
Abstract
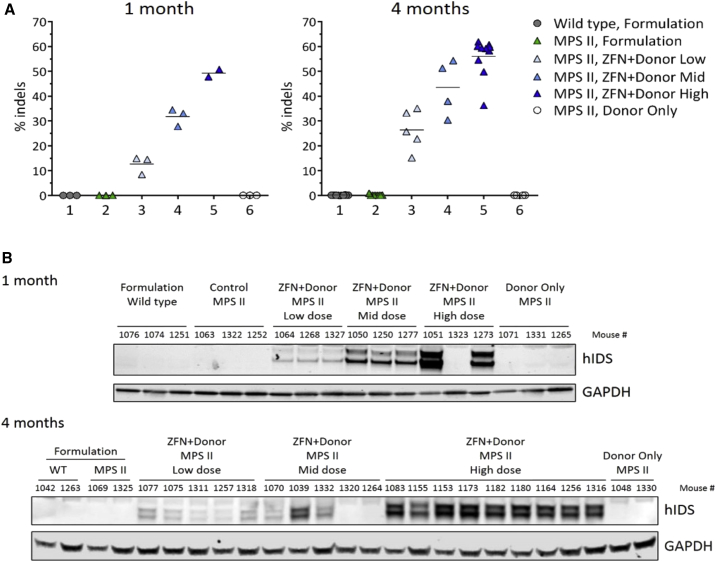
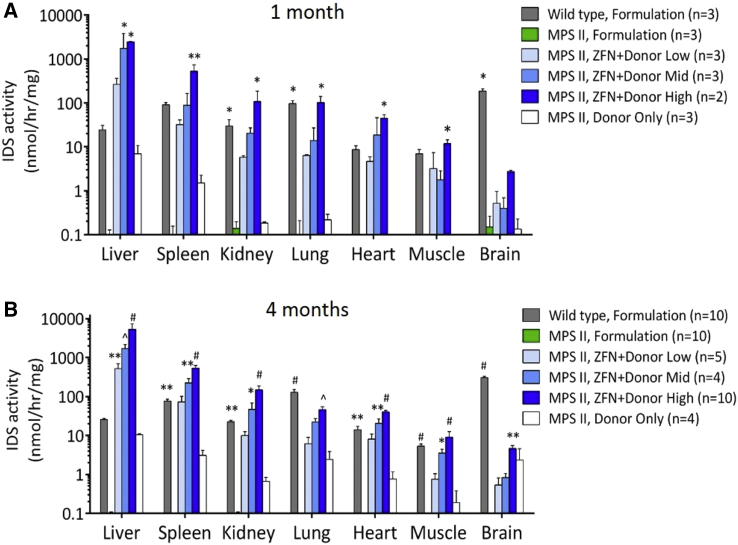
Mucopolysaccharidosis type II (MPS II) is an X-linked recessive lysosomal disorder caused by deficiency of iduronate 2-sulfatase (IDS), leading to accumulation of glycosaminoglycans (GAGs) in tissues of affected individuals, progressive disease, and shortened lifespan. Currently available enzyme replacement therapy (ERT) requires lifelong infusions and does not provide neurologic benefit. We utilized a zinc finger nuclease (ZFN)-targeting system to mediate genome editing for insertion of the human IDS (hIDS) coding sequence into a "safe harbor" site, intron 1 of the albumin locus in hepatocytes of an MPS II mouse model. Three dose levels of recombinant AAV2/8 vectors encoding a pair of ZFNs and a hIDS cDNA donor were administered systemically in MPS II mice. Supraphysiological, vector dose-dependent levels of IDS enzyme were observed in the circulation and peripheral organs of ZFN+donor-treated mice. GAG contents were markedly reduced in tissues from all ZFN+donor-treated groups. Surprisingly, we also demonstrate that ZFN-mediated genome editing prevented the development of neurocognitive deficit in young MPS II mice (6-9 weeks old) treated at high vector dose levels. We conclude that this ZFN-based platform for expression of therapeutic proteins from the albumin locus is a promising approach for treatment of MPS II and other lysosomal diseases.
Keywords: Hunter syndrome; MPS II; albumin locus; gene therapy; iduronate 2-sulfatase; in vivo genome editing; lysosomal disease; zinc finger nuclease.
Copyright © 2018 The Authors. Published by Elsevier Inc. All rights reserved.
Figures




References
-
- Lombardo A., Genovese P., Beausejour C.M., Colleoni S., Lee Y.-L., Kim K.A., Ando D., Urnov F.D., Galli C., Gregory P.D. Gene editing in human stem cells using zinc finger nucleases and integrase-defective lentiviral vector delivery. Nat. Biotechnol. 2007;25:1298–1306. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
