

Lévy flight movements prevent extinctions and maximize population abundances in fragile Lotka-Volterra systems
- PMID: 29581271
- PMCID: PMC5899458
- DOI: 10.1073/pnas.1719889115
Lévy flight movements prevent extinctions and maximize population abundances in fragile Lotka-Volterra systems
Abstract
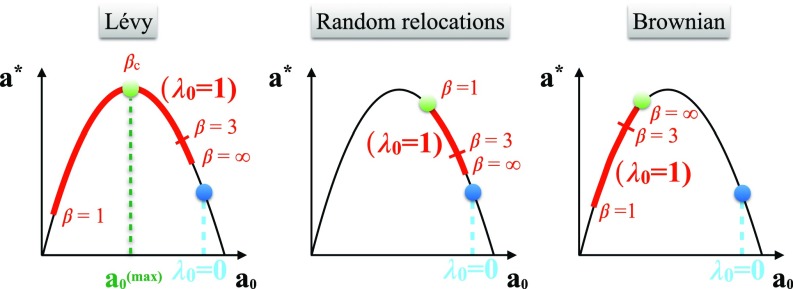
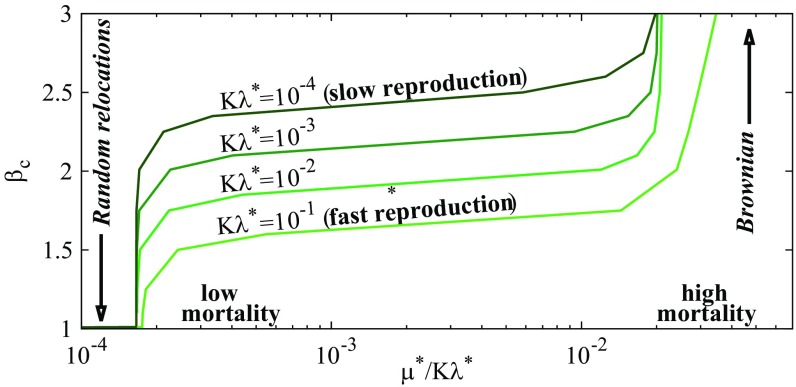
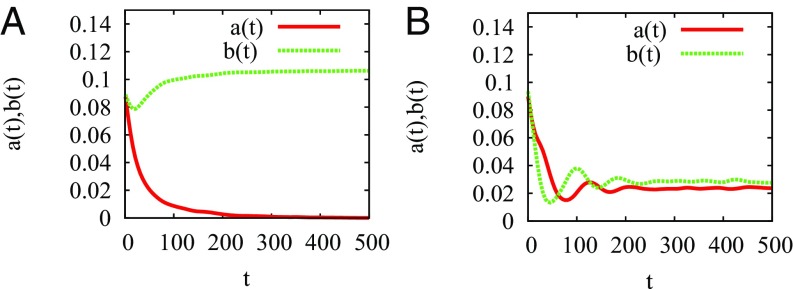
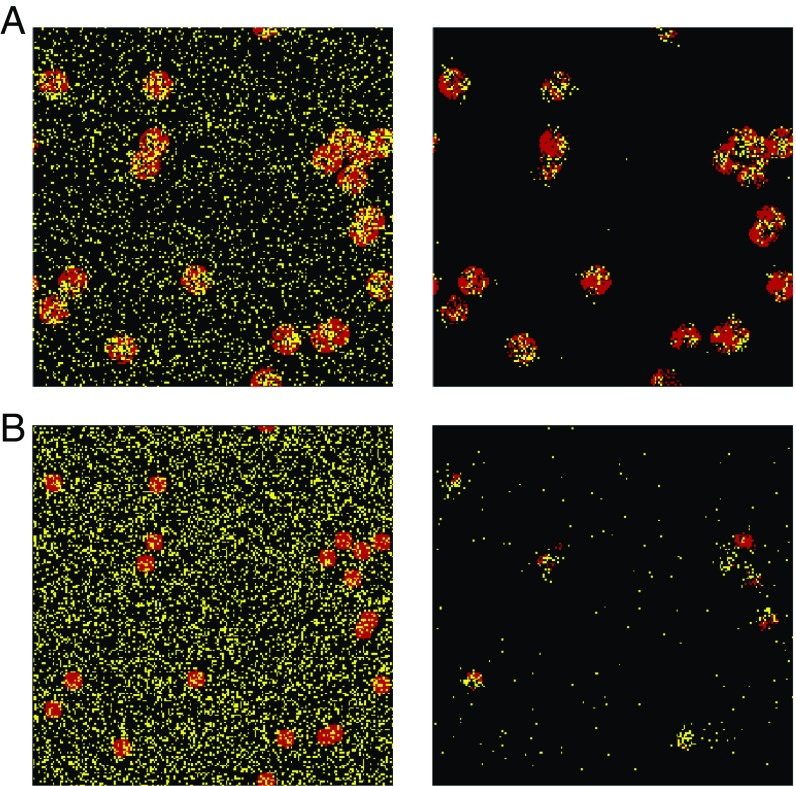
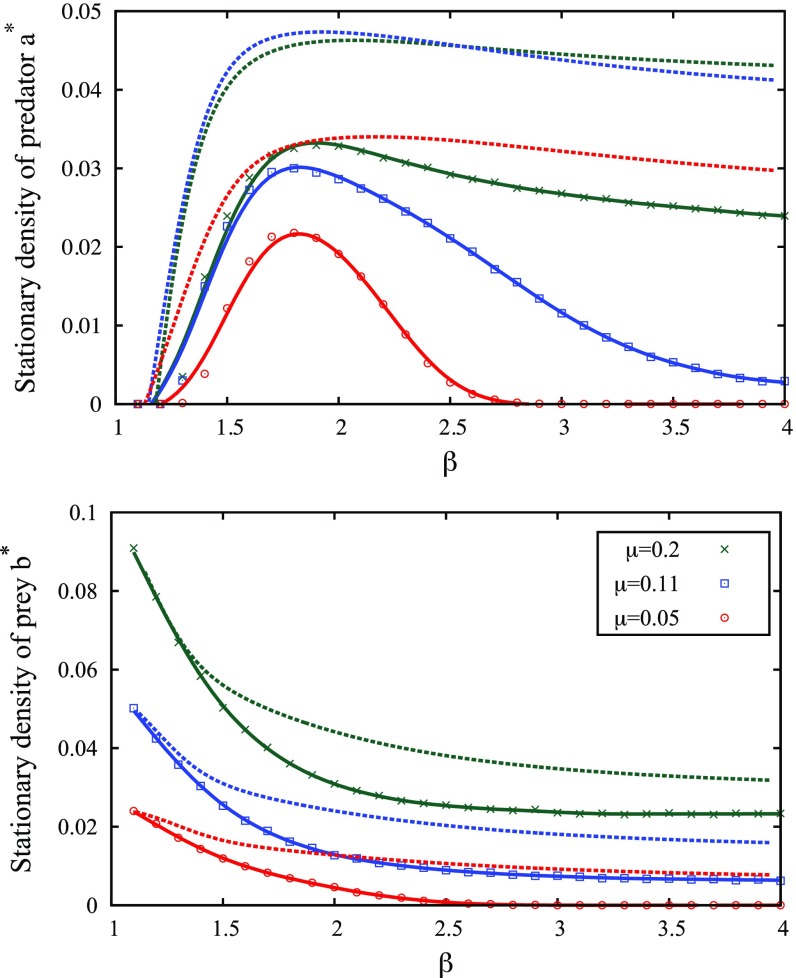
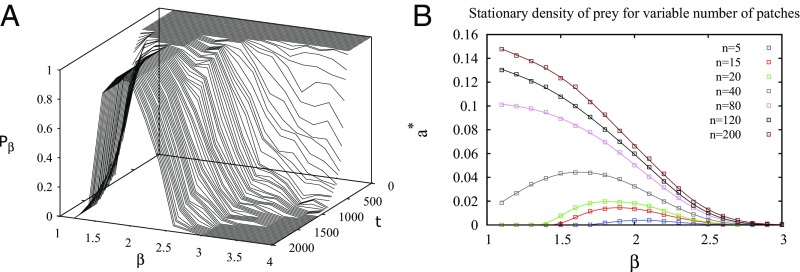
Multiple-scale mobility is ubiquitous in nature and has become instrumental for understanding and modeling animal foraging behavior. However, the impact of individual movements on the long-term stability of populations remains largely unexplored. We analyze deterministic and stochastic Lotka-Volterra systems, where mobile predators consume scarce resources (prey) confined in patches. In fragile systems (that is, those unfavorable to species coexistence), the predator species has a maximized abundance and is resilient to degraded prey conditions when individual mobility is multiple scaled. Within the Lévy flight model, highly superdiffusive foragers rarely encounter prey patches and go extinct, whereas normally diffusing foragers tend to proliferate within patches, causing extinctions by overexploitation. Lévy flights of intermediate index allow a sustainable balance between patch exploitation and regeneration over wide ranges of demographic rates. Our analytical and simulated results can explain field observations and suggest that scale-free random movements are an important mechanism by which entire populations adapt to scarcity in fragmented ecosystems.
Keywords: Lotka–Volterra; Lévy flights; ecological modeling; foraging; metapopulations.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Valiente-Banuet A, et al. Beyond species loss: The extinction of ecological interactions in a changing world. Funct Ecol. 2015;29:299–307.
-
- Barnosky AD, et al. Has the earth’s sixth mass extinction already arrived? Nature. 2011;471:51–57. - PubMed
-
- De Vos JM, Joppa LN, Gittleman JL, Stephens PR, Pimm SL. Estimating the normal background rate of species extinction. Conserv Biol. 2015;29:452–462. - PubMed
-
- May RM. Stability and Complexity in Model Ecosystems. Vol 6 Princeton Univ Press; Princeton: 2001.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
