

Atrophy of primary lymphoid organs induced by Marek's disease virus during early infection is associated with increased apoptosis, inhibition of cell proliferation and a severe B-lymphopenia
- PMID: 29587836
- PMCID: PMC5870490
- DOI: 10.1186/s13567-018-0526-x
Atrophy of primary lymphoid organs induced by Marek's disease virus during early infection is associated with increased apoptosis, inhibition of cell proliferation and a severe B-lymphopenia
Abstract
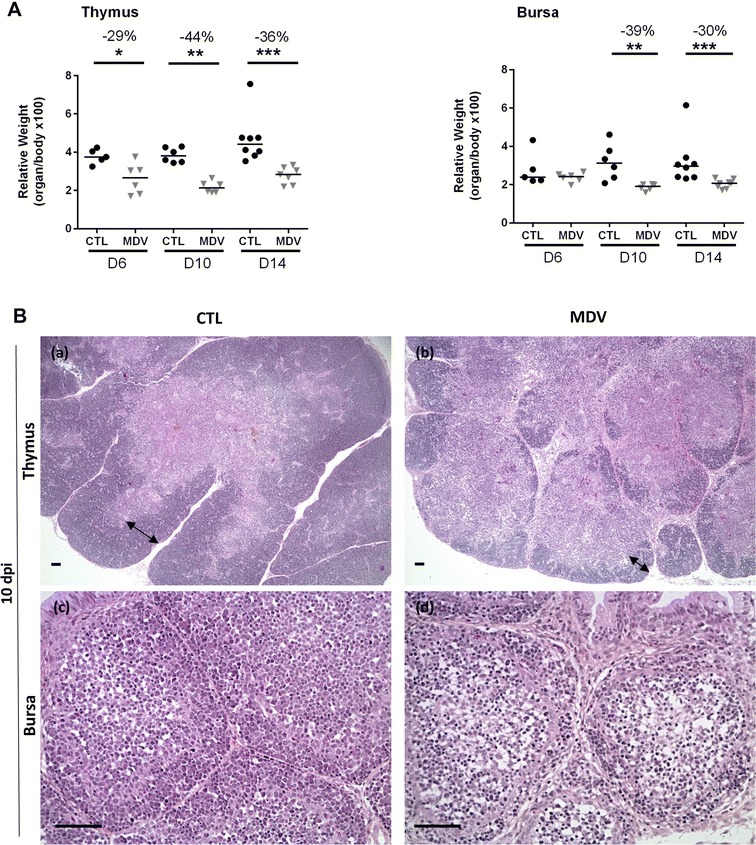
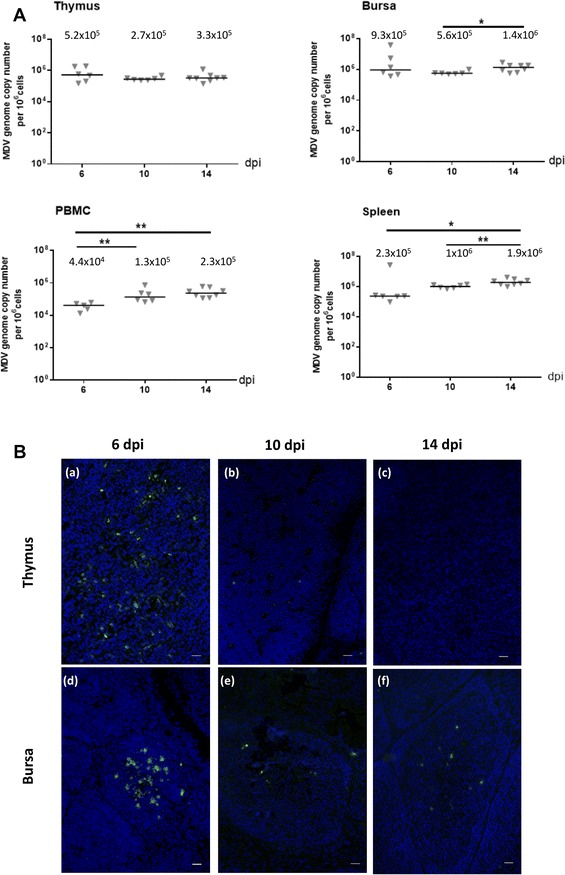
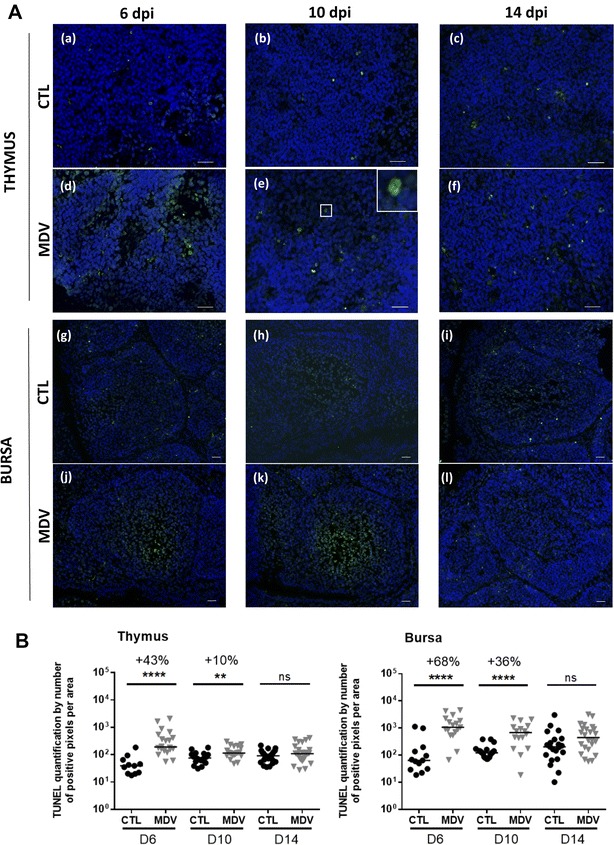
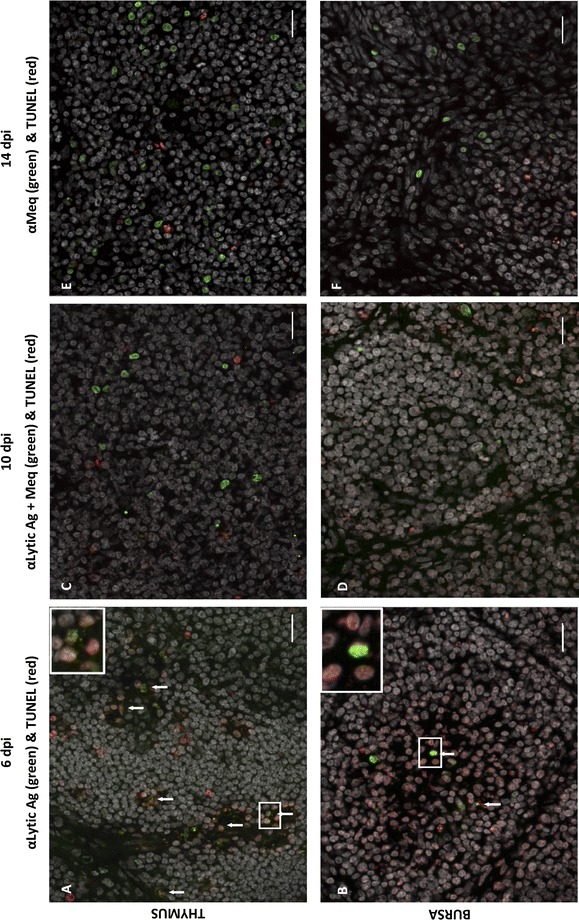
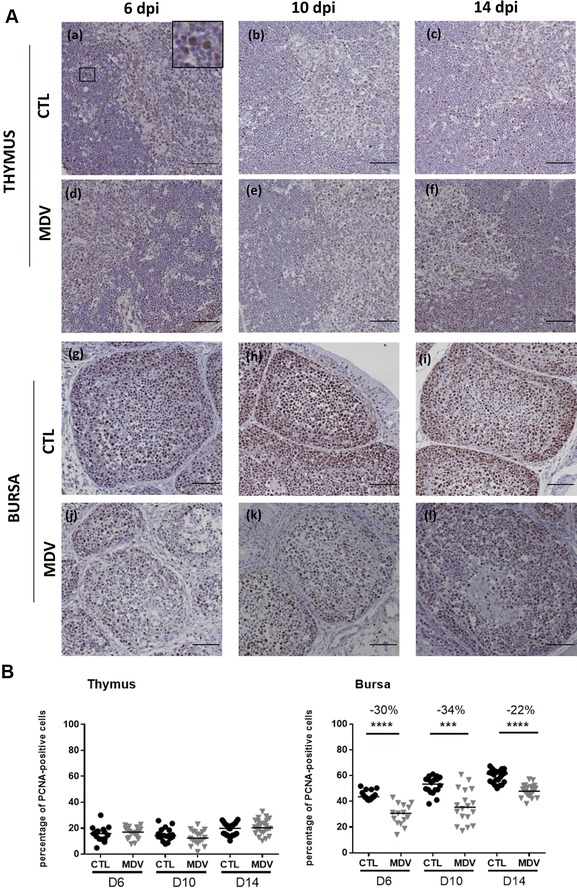
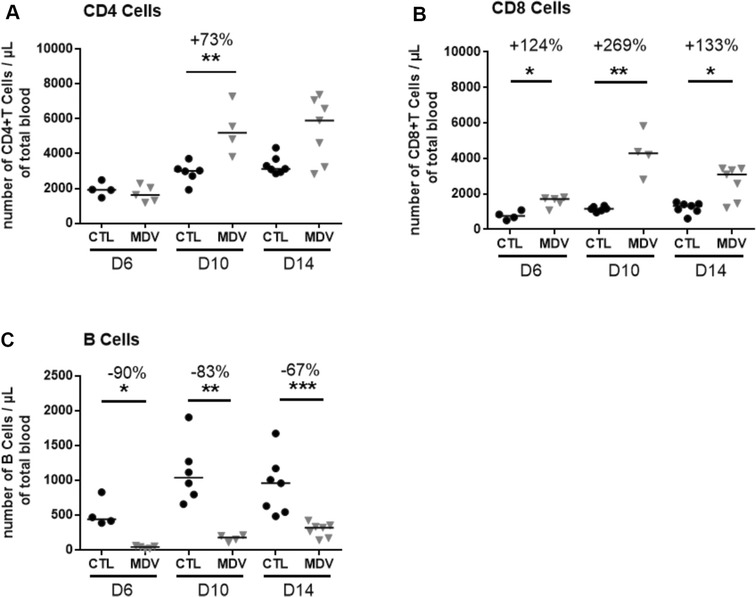
Marek's disease is a multi-faceted highly contagious disease affecting chickens caused by the Marek's disease alphaherpesvirus (MDV). MDV early infection induces a transient immunosuppression, which is associated with thymus and bursa of Fabricius atrophy. Little is known about the cellular processes involved in primary lymphoid organ atrophy. Here, by in situ TUNEL assay, we demonstrate that MDV infection results in a high level of apoptosis in the thymus and bursa of Fabricius, which is concomitant to the MDV lytic cycle. Interestingly, we observed that in the thymus most of the MDV infected cells at 6 days post-infection (dpi) were apoptotic, whereas in the bursa of Fabricius most of the apoptotic cells were uninfected suggesting that MDV triggers apoptosis by two different modes in these two primary lymphoid organs. In addition, a high decrease of cell proliferation was observed from 6 to 14 dpi in the bursa of Fabricius follicles, and not in the thymus. Finally, with an adapted absolute blood lymphocyte count, we demonstrate a major B-lymphopenia during the two 1st weeks of infection, and propose this method as a potent non-invasive tool to diagnose MDV bursa of Fabricius infection and atrophy. Our results demonstrate that the thymus and bursa of Fabricius atrophies are related to different cell mechanisms, with different temporalities, that affect infected and uninfected cells.
Figures
References
-
- Morrow C, Fehler F. Marek’s disease: a worldwide problem. In: Davison F, Nair V, editors. Marek’s disease: an evolving problem. London: Elsevier Academic Press; 2004. pp. 49–61.
-
- Calnek BW. Lymphomagenesis in Marek’s disease. Avian Pathol. 1998;27:S54–S64. doi: 10.1080/03079459808419293. - DOI
-
- Schat KA. Marek’s disease immunosuppression. In: Davison F, Nair V, editors. Marek’s disease: an evolving problem. London: Elsevier Academic Press; 2004. pp. 17–31.
-
- Schat KA, Nair V. Marek’s disease. In: Saif YM, editor. Disease of poultry. Ames: Blackwell Publishing Ltd; 2008. pp. 452–514.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
