

Microbial symbionts regulate the primary Ig repertoire
- PMID: 29588346
- PMCID: PMC5940265
- DOI: 10.1084/jem.20171761
Microbial symbionts regulate the primary Ig repertoire
Abstract
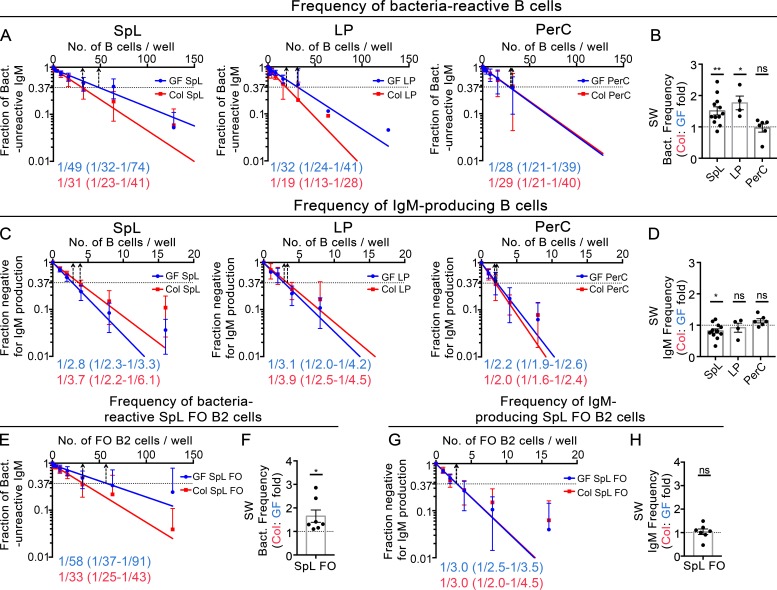
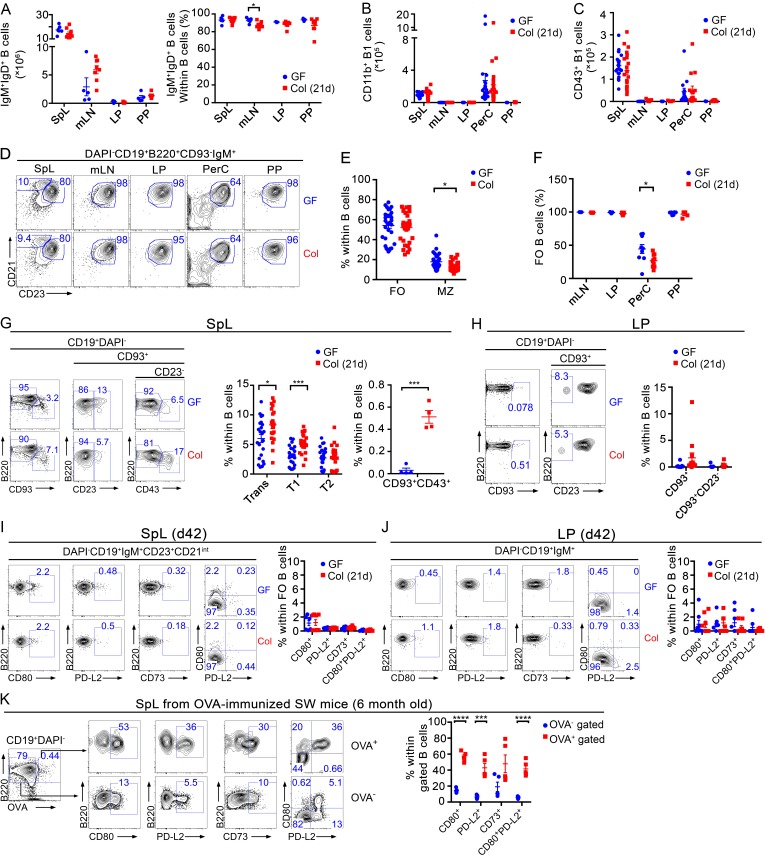
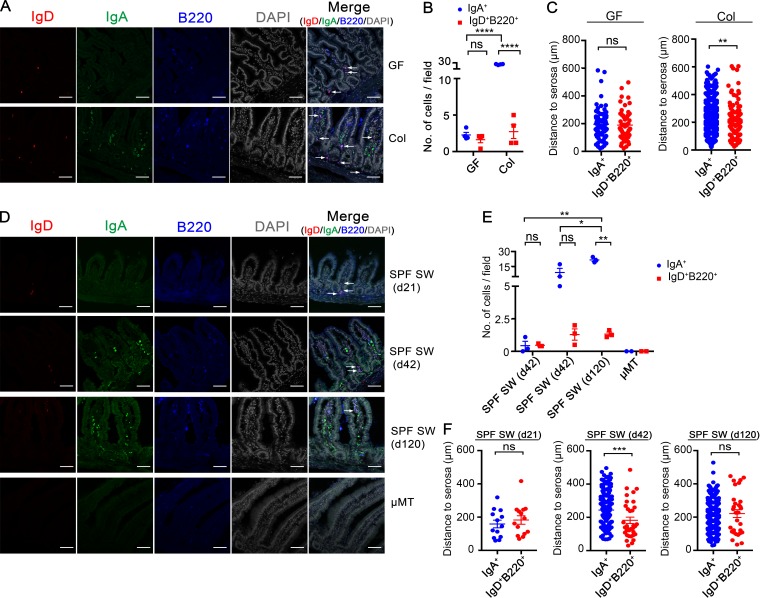
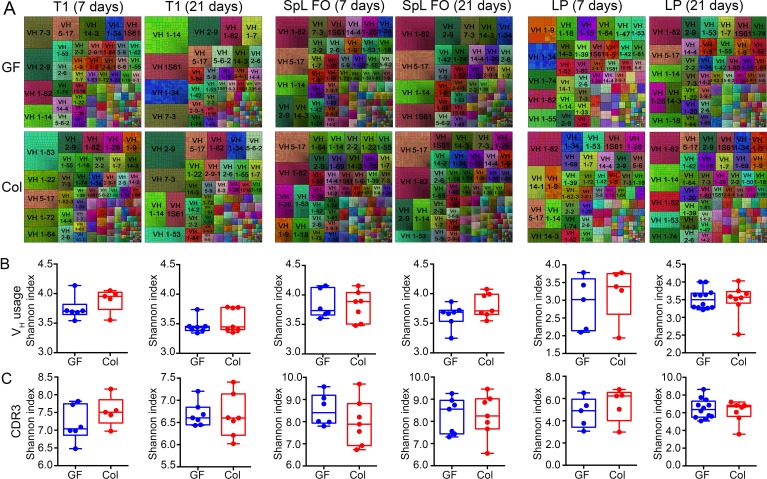
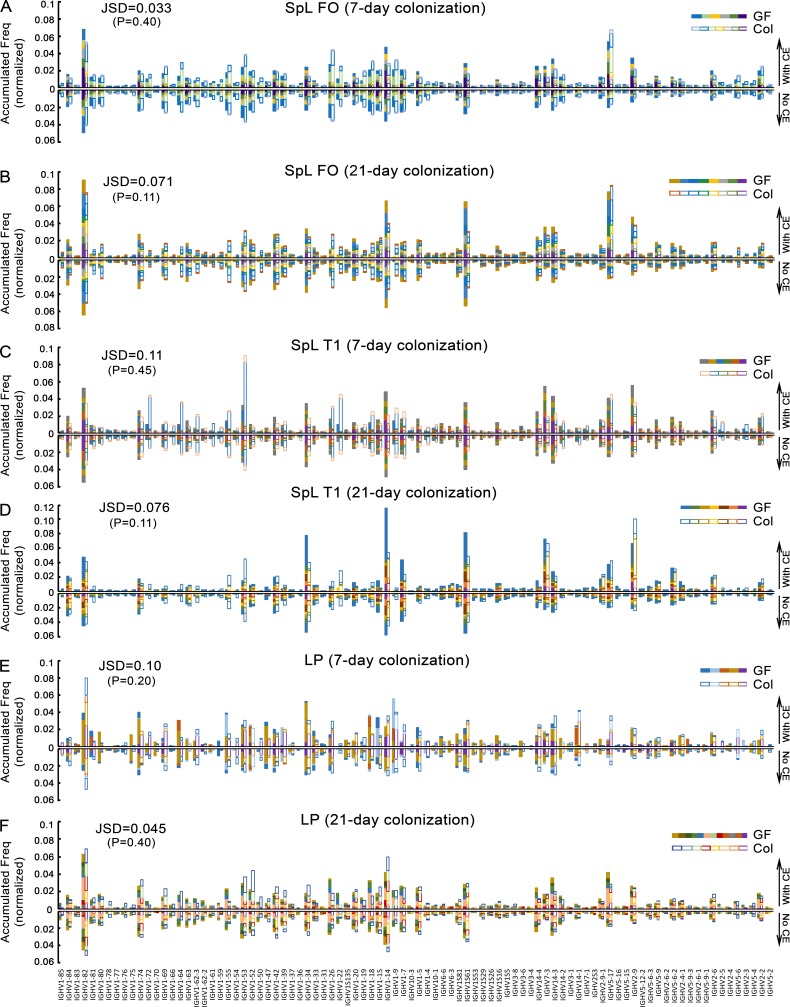
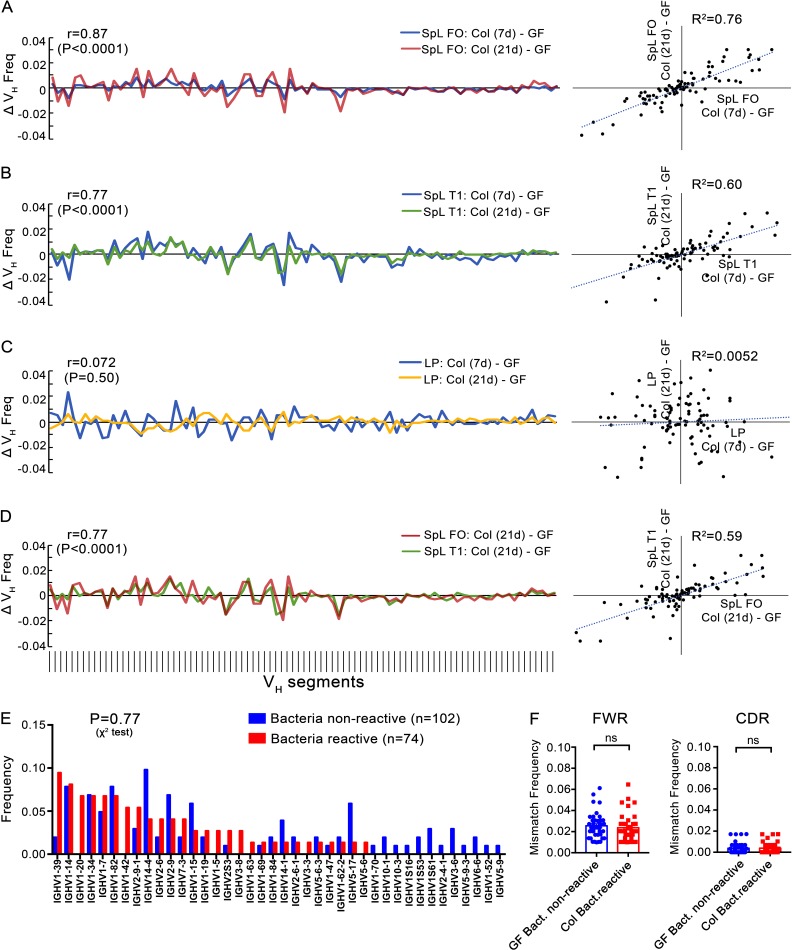
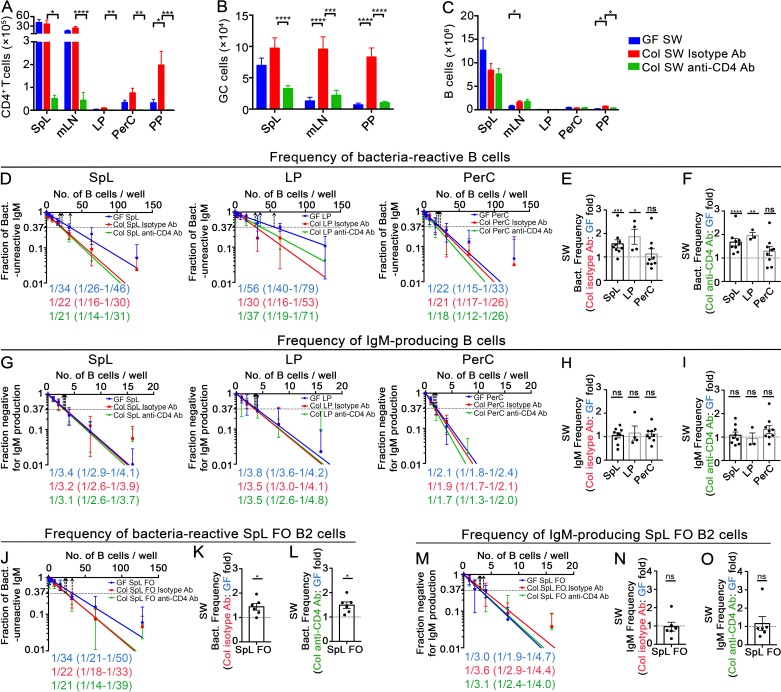
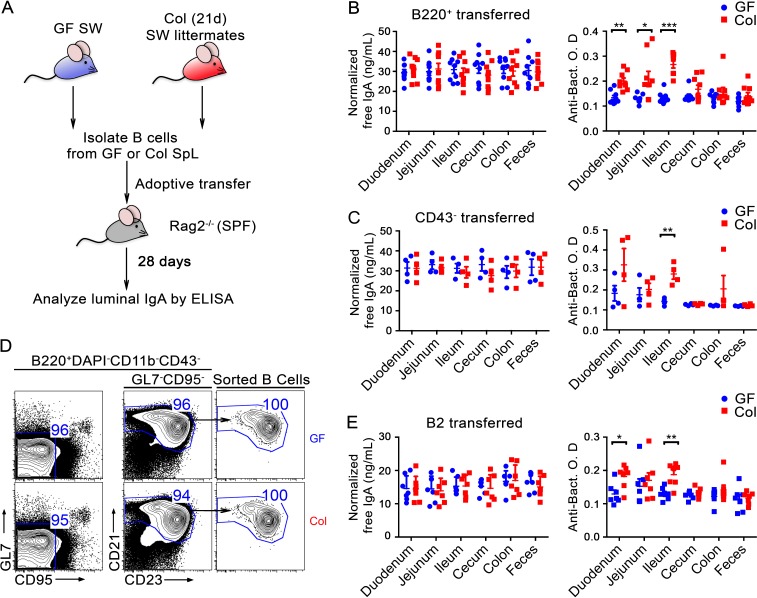
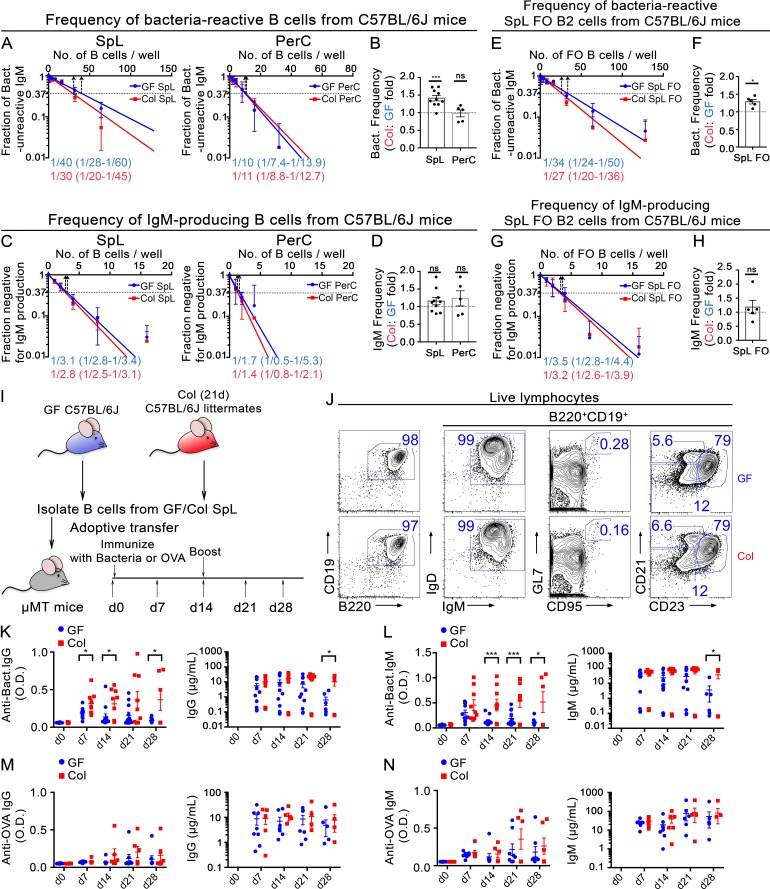
The ability of immunoglobulin (Ig) to recognize pathogens is critical for optimal immune fitness. Early events that shape preimmune Ig repertoires, expressed on IgM+ IgD+ B cells as B cell receptors (BCRs), are poorly defined. Here, we studied germ-free mice and conventionalized littermates to explore the hypothesis that symbiotic microbes help shape the preimmune Ig repertoire. Ig-binding assays showed that exposure to conventional microbial symbionts enriched frequencies of antibacterial IgM+ IgD+ B cells in intestine and spleen. This enrichment affected follicular B cells, involving a diverse set of Ig-variable region gene segments, and was T cell-independent. Functionally, enrichment of microbe reactivity primed basal levels of small intestinal T cell-independent, symbiont-reactive IgA and enhanced systemic IgG responses to bacterial immunization. These results demonstrate that microbial symbionts influence host immunity by enriching frequencies of antibacterial specificities within preimmune B cell repertoires and that this may have consequences for mucosal and systemic immunity.
© 2018 Chen et al.
Figures
References
-
- Bunker J.J., Flynn T.M., Koval J.C., Shaw D.G., Meisel M., McDonald B.D., Ishizuka I.E., Dent A.L., Wilson P.C., Jabri B., et al. 2015. Innate and Adaptive Humoral Responses Coat Distinct Commensal Bacteria with Immunoglobulin A. Immunity. 43:541–553. 10.1016/j.immuni.2015.08.007 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
