

Sequence-dependent catalytic regulation of the SpoIIIE motor activity ensures directionality of DNA translocation
- PMID: 29588476
- PMCID: PMC5869595
- DOI: 10.1038/s41598-018-23400-8
Sequence-dependent catalytic regulation of the SpoIIIE motor activity ensures directionality of DNA translocation
Abstract
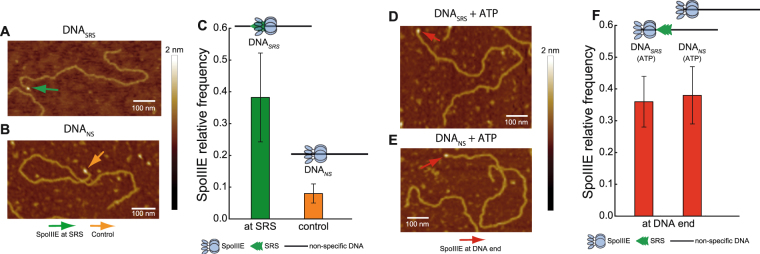
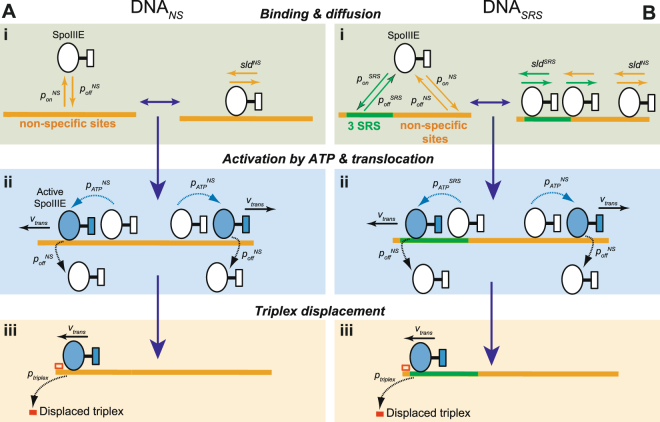
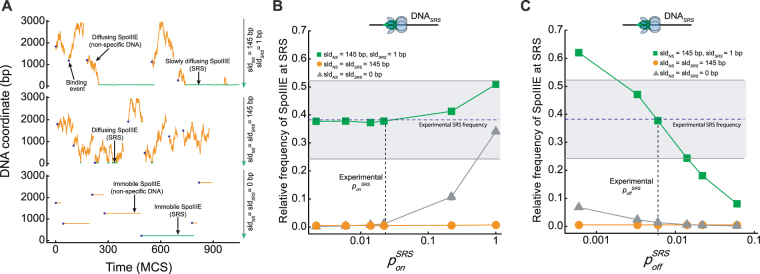
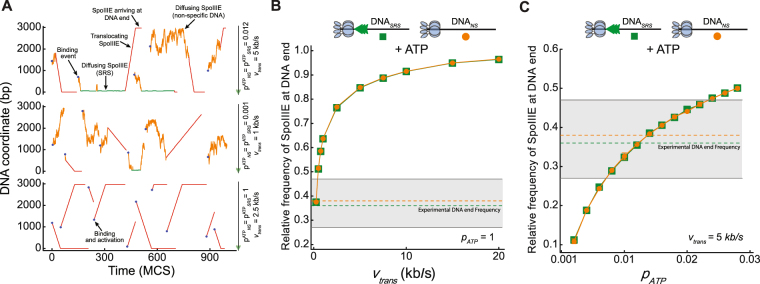
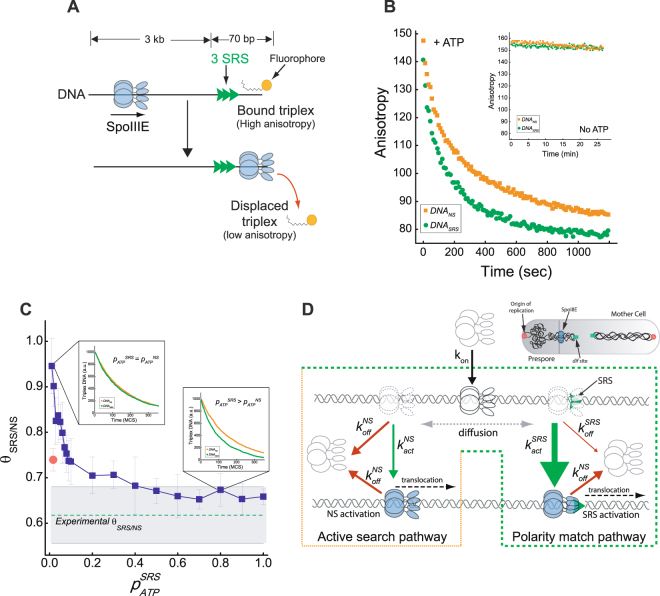
Transport of cellular cargo by molecular motors requires directionality to ensure proper biological functioning. During sporulation in Bacillus subtilis, directionality of chromosome transport is mediated by the interaction between the membrane-bound DNA translocase SpoIIIE and specific octameric sequences (SRS). Whether SRS regulate directionality by recruiting and orienting SpoIIIE or by simply catalyzing its translocation activity is still unclear. By using atomic force microscopy and single-round fast kinetics translocation assays we determined the localization and dynamics of diffusing and translocating SpoIIIE complexes on DNA with or without SRS. Our findings combined with mathematical modelling revealed that SpoIIIE directionality is not regulated by protein recruitment to SRS but rather by a fine-tuned balance among the rates governing SpoIIIE-DNA interactions and the probability of starting translocation modulated by SRS. Additionally, we found that SpoIIIE can start translocation from non-specific DNA, providing an alternative active search mechanism for SRS located beyond the exploratory length defined by 1D diffusion. These findings are relevant in vivo in the context of chromosome transport through an open channel, where SpoIIIE can rapidly explore DNA while directionality is modulated by the probability of translocation initiation upon interaction with SRS versus non-specific DNA.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
