

Molecularly barcoded Zika virus libraries to probe in vivo evolutionary dynamics
- PMID: 29590202
- PMCID: PMC5891079
- DOI: 10.1371/journal.ppat.1006964
Molecularly barcoded Zika virus libraries to probe in vivo evolutionary dynamics
Abstract
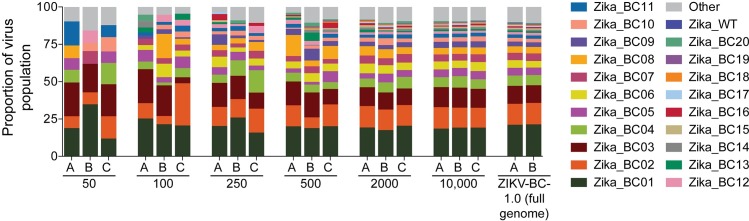
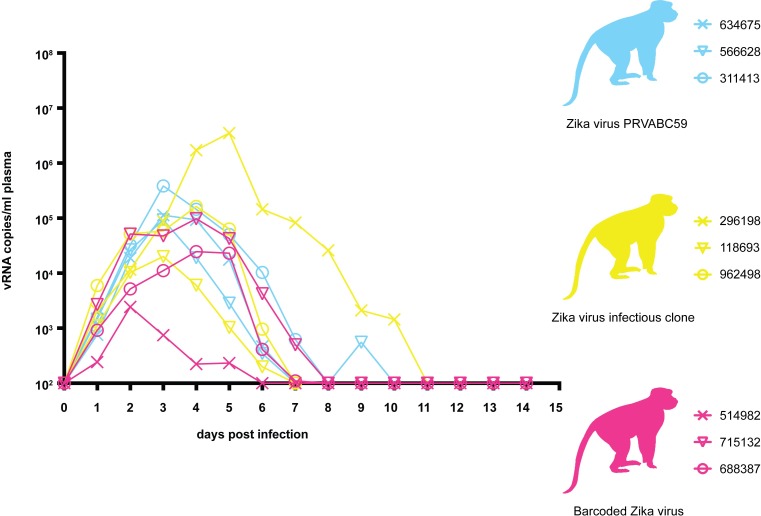
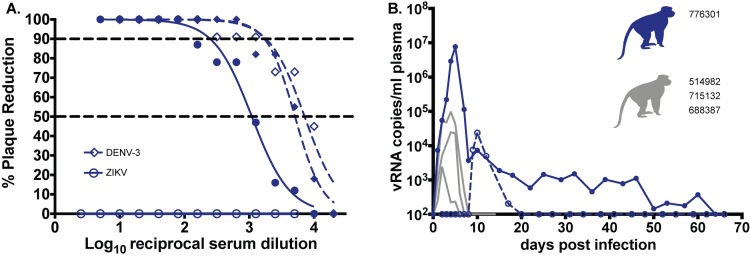
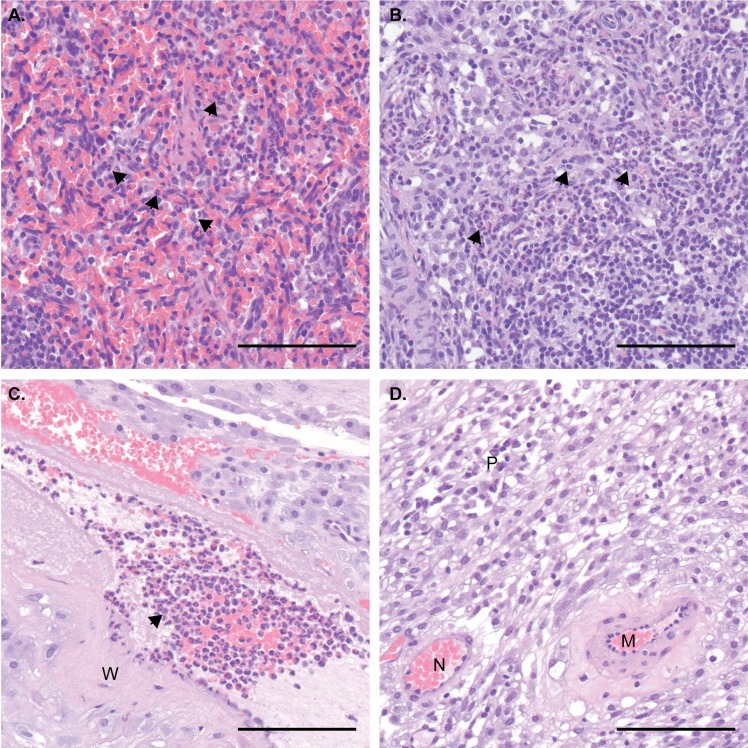
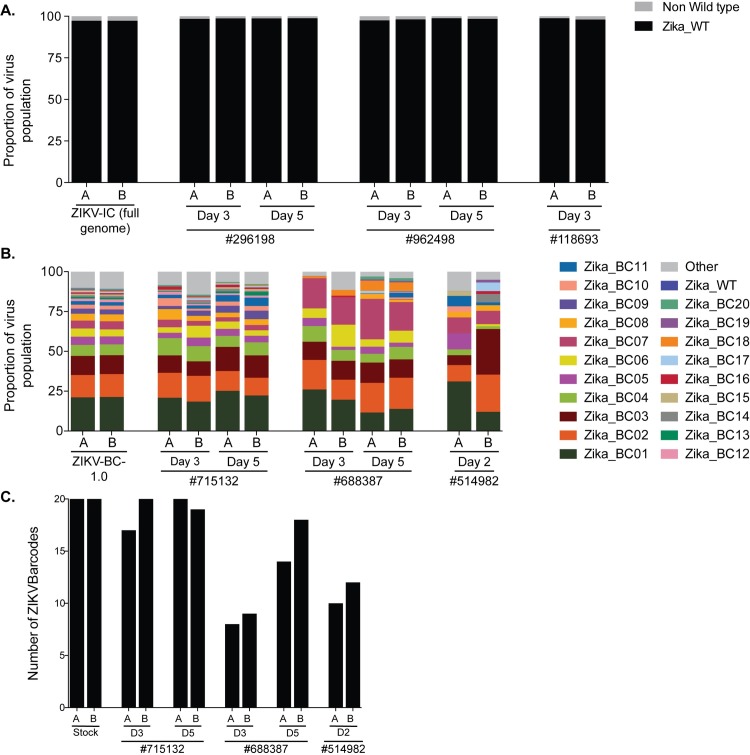
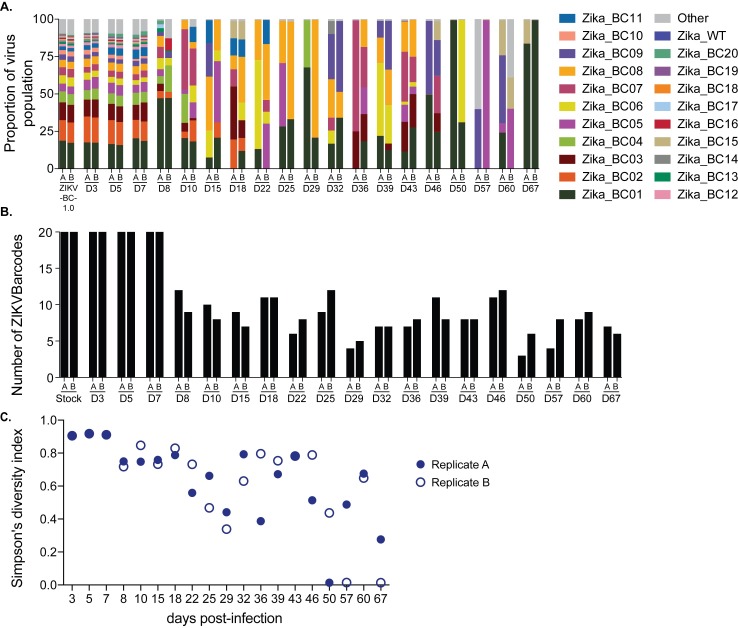
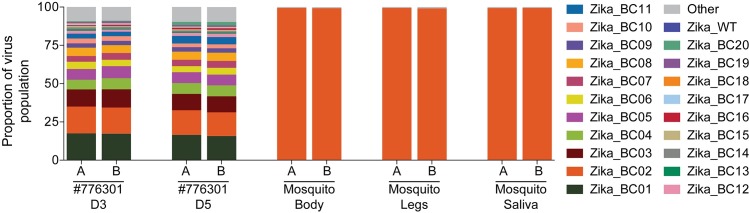
Defining the complex dynamics of Zika virus (ZIKV) infection in pregnancy and during transmission between vertebrate hosts and mosquito vectors is critical for a thorough understanding of viral transmission, pathogenesis, immune evasion, and potential reservoir establishment. Within-host viral diversity in ZIKV infection is low, which makes it difficult to evaluate infection dynamics. To overcome this biological hurdle, we constructed a molecularly barcoded ZIKV. This virus stock consists of a "synthetic swarm" whose members are genetically identical except for a run of eight consecutive degenerate codons, which creates approximately 64,000 theoretical nucleotide combinations that all encode the same amino acids. Deep sequencing this region of the ZIKV genome enables counting of individual barcodes to quantify the number and relative proportions of viral lineages present within a host. Here we used these molecularly barcoded ZIKV variants to study the dynamics of ZIKV infection in pregnant and non-pregnant macaques as well as during mosquito infection/transmission. The barcoded virus had no discernible fitness defects in vivo, and the proportions of individual barcoded virus templates remained stable throughout the duration of acute plasma viremia. ZIKV RNA also was detected in maternal plasma from a pregnant animal infected with barcoded virus for 67 days. The complexity of the virus population declined precipitously 8 days following infection of the dam, consistent with the timing of typical resolution of ZIKV in non-pregnant macaques and remained low for the subsequent duration of viremia. Our approach showed that synthetic swarm viruses can be used to probe the composition of ZIKV populations over time in vivo to understand vertical transmission, persistent reservoirs, bottlenecks, and evolutionary dynamics.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Reynolds MR, Jones AM, Petersen EE, Lee EH, Rice ME, Bingham A et al. (2017) Vital Signs: Update on Zika Virus-Associated Birth Defects and Evaluation of All U.S. Infants with Congenital Zika Virus Exposure—U.S. Zika Pregnancy Registry, 2016. MMWR Morb Mortal Wkly Rep 66: 366–373. doi: 10.15585/mmwr.mm6613e1 - DOI - PMC - PubMed
-
- Nguyen SM, Antony KM, Dudley DM, Kohn S, Simmons HA, Wolfe B et al. (2017) Highly efficient maternal-fetal Zika virus transmission in pregnant rhesus macaques. PLoS Pathog 13: e1006378 doi: 10.1371/journal.ppat.1006378 - DOI - PMC - PubMed
-
- Rosenberg K (2017) Zika Virus can Persist in Body Fluids for Prolonged Periods. Am J Nurs 117: 71. - PubMed
-
- Musso D, Roche C, Robin E, Nhan T, Teissier A, Cao-Lormeau VM (2015) Potential sexual transmission of Zika virus. Emerg Infect Dis 21: 359–361. doi: 10.3201/eid2102.141363 - DOI - PMC - PubMed
-
- Aid M, Abbink P, Larocca RA, Boyd M, Nityanandam R, Nanayakkara O et al. (2017) Zika Virus Persistence in the Central Nervous System and Lymph Nodes of Rhesus Monkeys. Cell 169: 610–620.e14. doi: 10.1016/j.cell.2017.04.008 - DOI - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
