

Entropic Control of Receptor Recycling Using Engineered Ligands
- PMID: 29590595
- PMCID: PMC5883623
- DOI: 10.1016/j.bpj.2018.01.036
Entropic Control of Receptor Recycling Using Engineered Ligands
Abstract
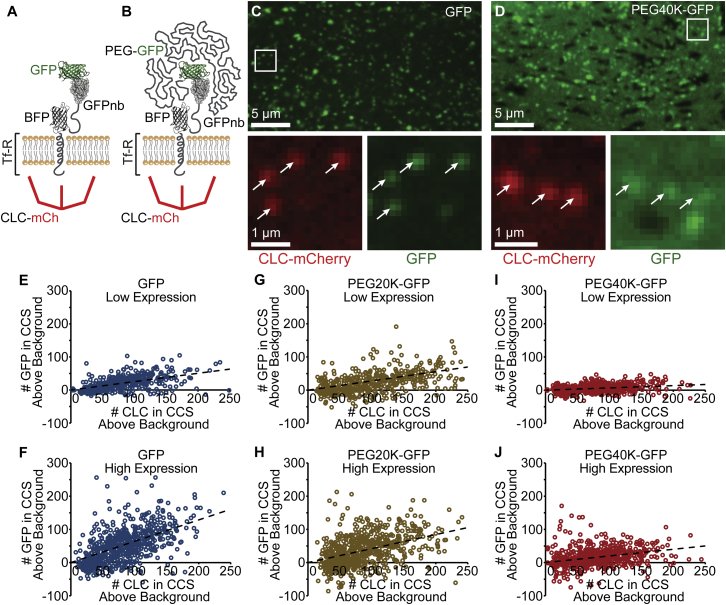
Receptor internalization by endocytosis regulates diverse cellular processes, from the rate of nutrient uptake to the timescale of essential signaling events. The established view is that internalization is tightly controlled by specific protein-binding interactions. However, recent work suggests that physical aspects of receptors influence the process in ways that cannot be explained by biochemistry alone. Specifically, work from several groups suggests that increasing the steric bulk of receptors may inhibit their uptake by multiple types of trafficking vesicles. How do biochemical and biophysical factors work together to control internalization? Here, we show that receptor uptake is well described by a thermodynamic trade-off between receptor-vesicle binding energy and the entropic cost of confining receptors within endocytic vesicles. Specifically, using large ligands to acutely increase the size of engineered variants of the transferrin receptor, we demonstrate that an increase in the steric bulk of a receptor dramatically decreases its probability of uptake by clathrin-coated structures. Further, in agreement with a simple thermodynamic analysis, all data collapse onto a single trend relating fractional occupancy of the endocytic structure to fractional occupancy of the surrounding plasma membrane, independent of receptor size. This fundamental scaling law provides a simple tool for predicting the impact of receptor expression level, steric bulk, and the size of endocytic structures on receptor uptake. More broadly, this work suggests that bulky ligands could be used to drive the accumulation of specific receptors at the plasma membrane surface, providing a biophysical tool for targeted modulation of signaling and metabolism from outside the cell.
Copyright © 2018 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures



Comment in
-
Bigger Isn't Always Better: Bulking Up Impedes Receptor Internalization.Biophys J. 2018 Mar 27;114(6):1255-1256. doi: 10.1016/j.bpj.2018.01.037. Biophys J. 2018. PMID: 29590581 Free PMC article. No abstract available.
References
-
- Davis C.G., Lehrman M.A., Goldstein J.L. The J.D. mutation in familial hypercholesterolemia: amino acid substitution in cytoplasmic domain impedes internalization of LDL receptors. Cell. 1986;45:15–24. - PubMed
-
- Vieira A.V., Lamaze C., Schmid S.L. Control of EGF receptor signaling by clathrin-mediated endocytosis. Science. 1996;274:2086–2089. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
