

A Highly Conserved Leucine in Mammarenavirus Matrix Z Protein Is Required for Z Interaction with the Virus L Polymerase and Z Stability in Cells Harboring an Active Viral Ribonucleoprotein
- PMID: 29593035
- PMCID: PMC5952140
- DOI: 10.1128/JVI.02256-17
A Highly Conserved Leucine in Mammarenavirus Matrix Z Protein Is Required for Z Interaction with the Virus L Polymerase and Z Stability in Cells Harboring an Active Viral Ribonucleoprotein
Abstract
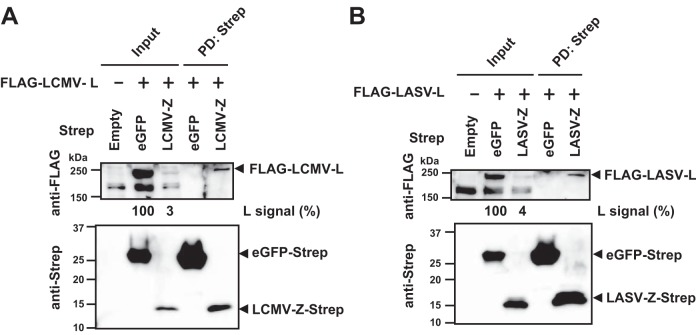
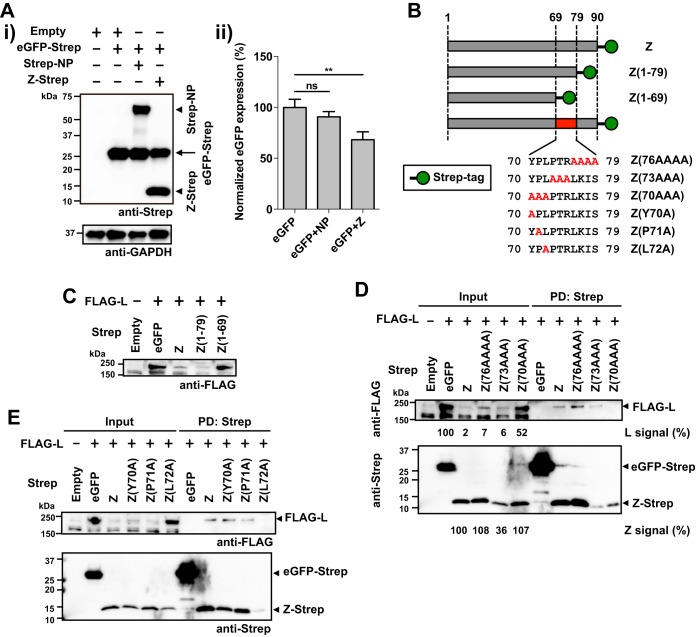
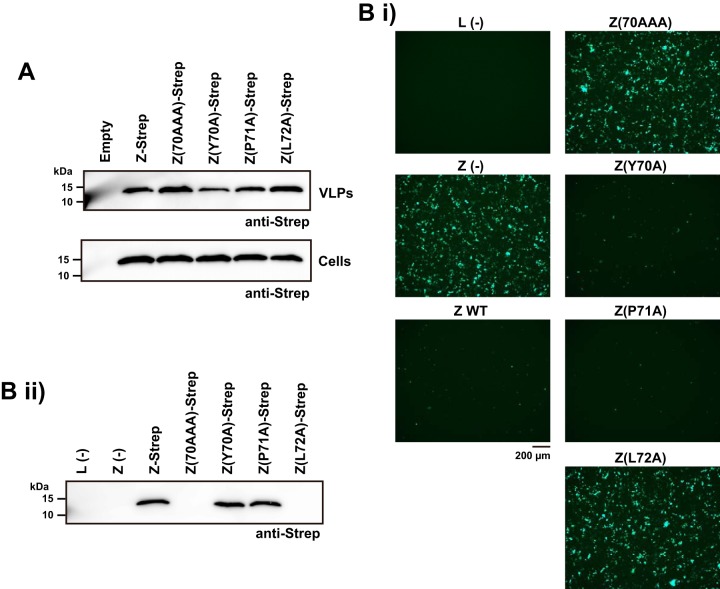
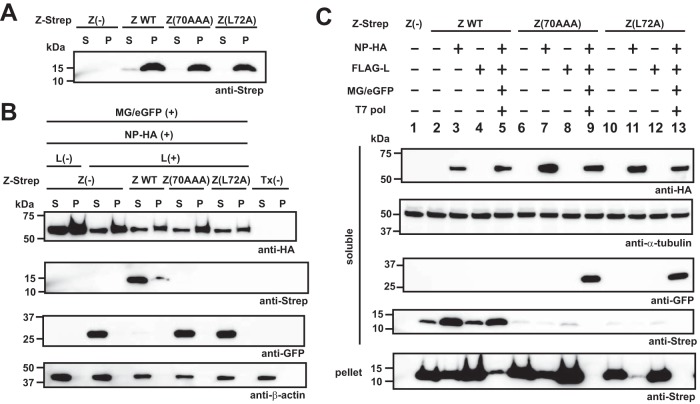
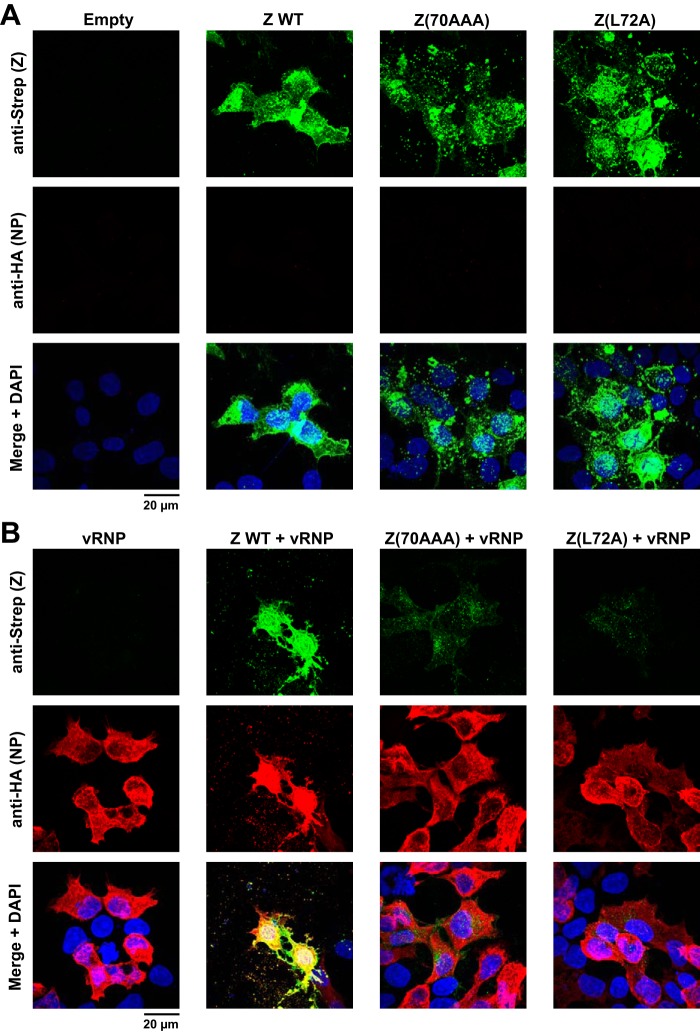
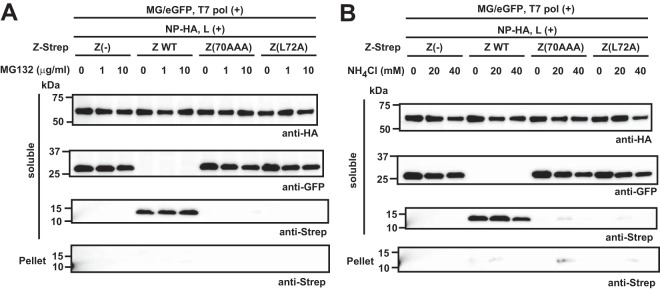
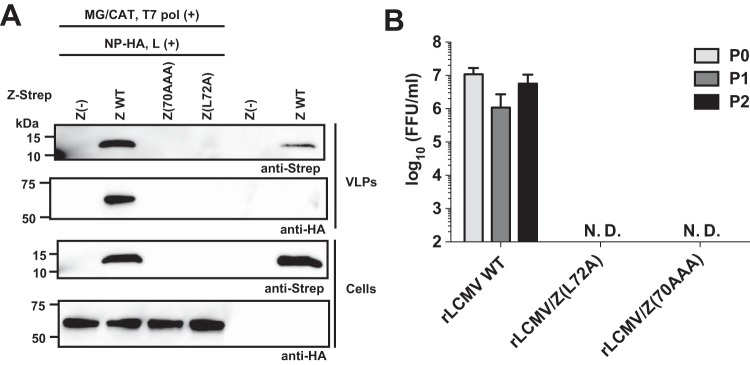
Mammarenaviruses cause chronic infections in their natural rodent hosts. Infected rodents shed infectious virus into excreta. Humans are infected through mucosal exposure to aerosols or direct contact of abraded skin with fomites, resulting in a wide range of manifestations from asymptomatic or mild febrile illness to severe life-threatening hemorrhagic fever. The mammarenavirus matrix Z protein has been shown to be a main driving force of virus budding and to act as a negative regulator of viral RNA synthesis. To gain a better understanding of how the Z protein exerts its several different functions, we investigated the interaction between Z and viral polymerase L protein using the prototypic mammarenavirus, lymphocytic choriomeningitis virus (LCMV). We found that in the presence of an active viral ribonucleoprotein (vRNP), the Z protein translocated from nonionic detergent-resistant, membrane-rich structures to a subcellular compartment with a different membrane composition susceptible to disruption by nonionic detergents. Alanine (A) substitution of a highly conserved leucine (L) at position 72 in LCMV Z protein abrogated Z-L interaction. The L72A mutation did not affect the stability or budding activity of Z when expressed alone, but in the presence of an active vRNP, mutation L72A promoted rapid degradation of Z via a proteasome- and lysosome-independent pathway. Accordingly, L72A mutation in the Z protein resulted in nonviable LCMV. Our findings have uncovered novel aspects of the dynamics of the Z protein for which a highly conserved L residue was strictly required.IMPORTANCE Several mammarenaviruses, chiefly Lassa virus (LASV), cause hemorrhagic fever disease in humans and pose important public health concerns in their regions of endemicity. Moreover, mounting evidence indicates that the worldwide-distributed, prototypic mammarenavirus, lymphocytic choriomeningitis virus (LCMV), is a neglected human pathogen of clinical significance. The mammarenavirus matrix Z protein plays critical roles in different steps of the viral life cycle by interacting with viral and host cellular components. Here we report that alanine substitution of a highly conserved leucine residue, located at position 72 in LCMV Z protein, abrogated Z-L interaction. The L72A mutation did not affect Z budding activity but promoted its rapid degradation in the presence of an active viral ribonucleoprotein (vRNP). Our findings have uncovered novel aspects of the dynamics of the Z protein for which a highly conserved L residue was strictly required.
Keywords: RNA-dependent RNA polymerase; arenavirus; matrix protein; protein stability.
Copyright © 2018 American Society for Microbiology.
Figures
Similar articles
-
Residues K465 and G467 within the Cytoplasmic Domain of GP2 Play a Critical Role in the Persistence of Lymphocytic Choriomeningitis Virus in Mice.J Virol. 2016 Oct 28;90(22):10102-10112. doi: 10.1128/JVI.01303-16. Print 2016 Nov 15. J Virol. 2016. PMID: 27581982 Free PMC article.
-
The small RING finger protein Z drives arenavirus budding: implications for antiviral strategies.Proc Natl Acad Sci U S A. 2003 Oct 28;100(22):12978-83. doi: 10.1073/pnas.2133782100. Epub 2003 Oct 16. Proc Natl Acad Sci U S A. 2003. PMID: 14563923 Free PMC article.
-
The C-terminal region of lymphocytic choriomeningitis virus nucleoprotein contains distinct and segregable functional domains involved in NP-Z interaction and counteraction of the type I interferon response.J Virol. 2011 Dec;85(24):13038-48. doi: 10.1128/JVI.05834-11. Epub 2011 Oct 5. J Virol. 2011. PMID: 21976642 Free PMC article.
-
Mammarenavirus Genetic Diversity and Its Biological Implications.Curr Top Microbiol Immunol. 2023;439:265-303. doi: 10.1007/978-3-031-15640-3_8. Curr Top Microbiol Immunol. 2023. PMID: 36592249 Review.
-
Lymphocytic choriomeningitis virus (LCMV) infection of macaques: a model for Lassa fever.Antiviral Res. 2011 Nov;92(2):125-38. doi: 10.1016/j.antiviral.2011.07.015. Epub 2011 Jul 27. Antiviral Res. 2011. PMID: 21820469 Free PMC article. Review.
Cited by
-
Isolation of Reconstructed Functional Ribonucleoprotein Complexes of Machupo Virus.J Virol. 2021 Oct 27;95(22):e0105421. doi: 10.1128/JVI.01054-21. Epub 2021 Aug 25. J Virol. 2021. PMID: 34432522 Free PMC article.
-
Mammarenavirus Z Protein Myristoylation and Oligomerization Are Not Required for Its Dose-Dependent Inhibitory Effect on vRNP Activity.Biochem (Basel). 2025 Jun;5(2):10. doi: 10.3390/biochem5020010. Epub 2025 Apr 29. Biochem (Basel). 2025. PMID: 40520408 Free PMC article.
-
Activation of the STAT3 Signaling Pathway by the RNA-Dependent RNA Polymerase Protein of Arenavirus.Viruses. 2021 May 25;13(6):976. doi: 10.3390/v13060976. Viruses. 2021. PMID: 34070281 Free PMC article.
-
Hemorrhagic Fever-Causing Arenaviruses: Lethal Pathogens and Potent Immune Suppressors.Front Immunol. 2019 Mar 13;10:372. doi: 10.3389/fimmu.2019.00372. eCollection 2019. Front Immunol. 2019. PMID: 30918506 Free PMC article. Review.
-
Characterization of bi-segmented and tri-segmented recombinant Pichinde virus particles.J Virol. 2024 Oct 22;98(10):e0079924. doi: 10.1128/jvi.00799-24. Epub 2024 Sep 12. J Virol. 2024. PMID: 39264155 Free PMC article.
References
-
- Buchmeier MJ, Peters CJ, de la Torre JC. 2007. Arenaviridae: the viruses and their replication, p 1791–1851. In Knipe DM, Howley PM, Griffin DE, Lamb RA, Martin MA, Roizman B, Straus SE (ed), Fields virology, 5th ed, vol 2 Lippincott Williams & Wilkins, Philadelphia, PA.
-
- Khan SH, Goba A, Chu M, Roth C, Healing T, Marx A, Fair J, Guttieri MC, Ferro P, Imes T, Monagin C, Garry RF, Bausch DG, Mano River Union Lassa Fever Network. 2008. New opportunities for field research on the pathogenesis and treatment of Lassa fever. Antiviral Res 78:103–115. doi:10.1016/j.antiviral.2007.11.003. - DOI - PubMed
-
- McCormick JB, Fisher-Hock SP. 2002. Lassa fever. Curr Top Microbiol Immunol 262:75–110. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
