

The early mature part of bacterial twin-arginine translocation (Tat) precursor proteins contributes to TatBC receptor binding
- PMID: 29593092
- PMCID: PMC5949997
- DOI: 10.1074/jbc.RA118.002576
The early mature part of bacterial twin-arginine translocation (Tat) precursor proteins contributes to TatBC receptor binding
Abstract
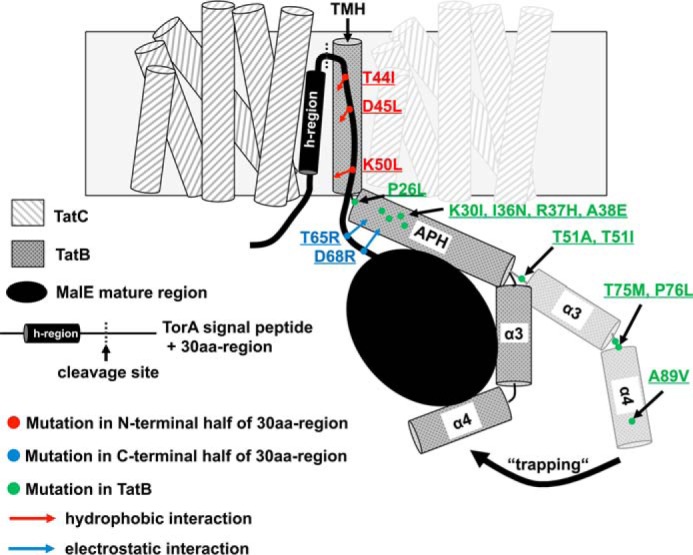
The twin-arginine translocation (Tat) pathway transports folded proteins across bacterial membranes. Tat precursor proteins possess a conserved twin-arginine (RR) motif in their signal peptides that is involved in the binding of the proteins to the membrane-associated TatBC receptor complex. In addition, the hydrophobic region in the Tat signal peptides also contributes to TatBC binding, but whether regions beyond the signal-peptide cleavage site are involved in this process is unknown. Here, we analyzed the contribution of the early mature protein part of the Escherichia coli trimethylamine N-oxide reductase (TorA) to productive TatBC receptor binding. We identified substitutions in the 30 amino acids immediately following the TorA signal peptide (30aa-region) that restored export of a transport-defective TorA[KQ]-30aa-MalE precursor, in which the RR residues had been replaced by a lysine-glutamine pair. Some of these substitutions increased the hydrophobicity of the N-terminal part of the 30aa-region and thereby likely enhanced hydrophobic substrate-receptor interactions within the hydrophobic TatBC substrate-binding cavity. Another class of substitutions increased the positive net charge of the region's C-terminal part, presumably leading to strengthened electrostatic interactions between the mature substrate part and the cytoplasmic TatBC regions. Furthermore, we identified substitutions in the C-terminal domains of TatB following the transmembrane segment that restored transport of various transport-defective TorA-MalE derivatives. Some of these substitutions most likely affected the orientation or conformation of the flexible, carboxy-proximal helices of TatB. Therefore, we propose that a tight accommodation of the folded mature region by TatB contributes to productive binding of Tat substrates to TatBC.
Keywords: Escherichia coli (E. coli); early mature region; membrane transport; protein export; protein folding; protein targeting; protein translocation; secretion pathway; substrate receptor; twin arginine translocation.
© 2018 Ulfig and Freudl.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article.
Figures




Similar articles
-
Targeting of proteins to the twin-arginine translocation pathway.Mol Microbiol. 2020 May;113(5):861-871. doi: 10.1111/mmi.14461. Epub 2020 Feb 20. Mol Microbiol. 2020. PMID: 31971282 Free PMC article. Review.
-
The h-region of twin-arginine signal peptides supports productive binding of bacterial Tat precursor proteins to the TatBC receptor complex.J Biol Chem. 2017 Jun 30;292(26):10865-10882. doi: 10.1074/jbc.M117.788950. Epub 2017 May 17. J Biol Chem. 2017. PMID: 28515319 Free PMC article.
-
Signal Peptide Hydrophobicity Modulates Interaction with the Twin-Arginine Translocase.mBio. 2017 Aug 1;8(4):e00909-17. doi: 10.1128/mBio.00909-17. mBio. 2017. PMID: 28765221 Free PMC article.
-
A signal sequence suppressor mutant that stabilizes an assembled state of the twin arginine translocase.Proc Natl Acad Sci U S A. 2017 Mar 7;114(10):E1958-E1967. doi: 10.1073/pnas.1615056114. Epub 2017 Feb 21. Proc Natl Acad Sci U S A. 2017. PMID: 28223511 Free PMC article.
-
Twin-arginine-dependent translocation of folded proteins.Philos Trans R Soc Lond B Biol Sci. 2012 Apr 19;367(1592):1029-46. doi: 10.1098/rstb.2011.0202. Philos Trans R Soc Lond B Biol Sci. 2012. PMID: 22411976 Free PMC article. Review.
Cited by
-
Targeting of proteins to the twin-arginine translocation pathway.Mol Microbiol. 2020 May;113(5):861-871. doi: 10.1111/mmi.14461. Epub 2020 Feb 20. Mol Microbiol. 2020. PMID: 31971282 Free PMC article. Review.
-
Analysis of the Brucella suis Twin Arginine Translocation System and Its Substrates Shows That It Is Essential for Viability.Infect Immun. 2023 Jan 24;91(1):e0045922. doi: 10.1128/iai.00459-22. Epub 2022 Nov 30. Infect Immun. 2023. PMID: 36448838 Free PMC article.
-
Twin-arginine translocase component TatB performs folding quality control via a chaperone-like activity.Sci Rep. 2022 Sep 1;12(1):14862. doi: 10.1038/s41598-022-18958-3. Sci Rep. 2022. PMID: 36050356 Free PMC article.
-
Surface-exposed domains of TatB involved in the structural and functional assembly of the Tat translocase in Escherichia coli.J Biol Chem. 2019 Sep 20;294(38):13902-13914. doi: 10.1074/jbc.RA119.009298. Epub 2019 Jul 24. J Biol Chem. 2019. PMID: 31341014 Free PMC article.
-
Synergistic Engineering of the Twin-Arginine Translocation (Tat) Pathway and Membrane Capacity Enhances Extracellular Production of Amylosucrase in Bacillus licheniformis.Microorganisms. 2025 May 22;13(6):1179. doi: 10.3390/microorganisms13061179. Microorganisms. 2025. PMID: 40572067 Free PMC article.
References
-
- Hou B., and Brüser T. (2011) The Tat-dependent protein translocation pathway. Biomol. Concepts 2, 507–523 - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
