

Vitamin C promotes apoptosis in breast cancer cells by increasing TRAIL expression
- PMID: 29593282
- PMCID: PMC5871772
- DOI: 10.1038/s41598-018-23714-7
Vitamin C promotes apoptosis in breast cancer cells by increasing TRAIL expression
Abstract
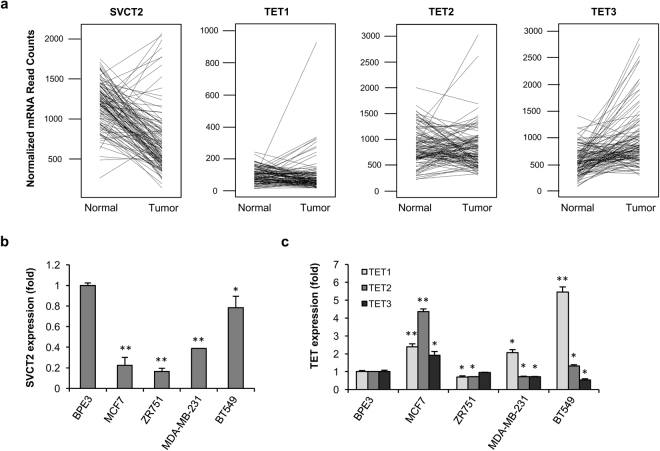
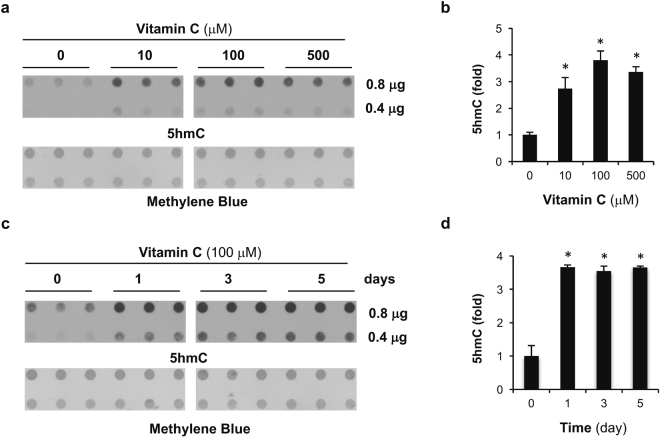
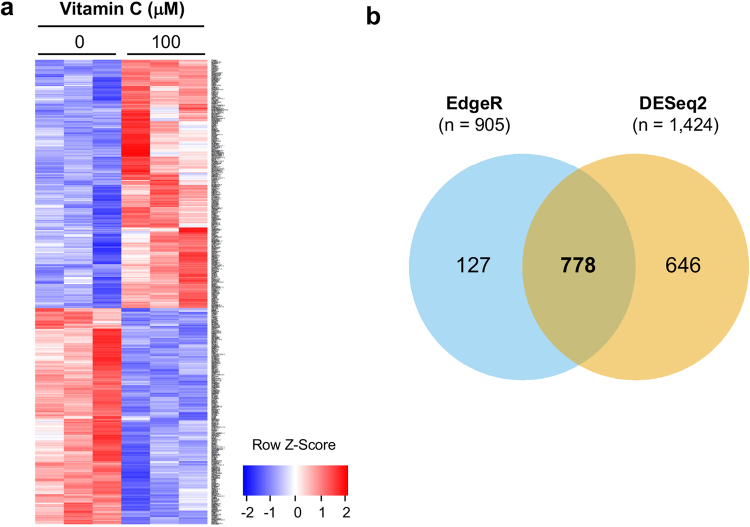
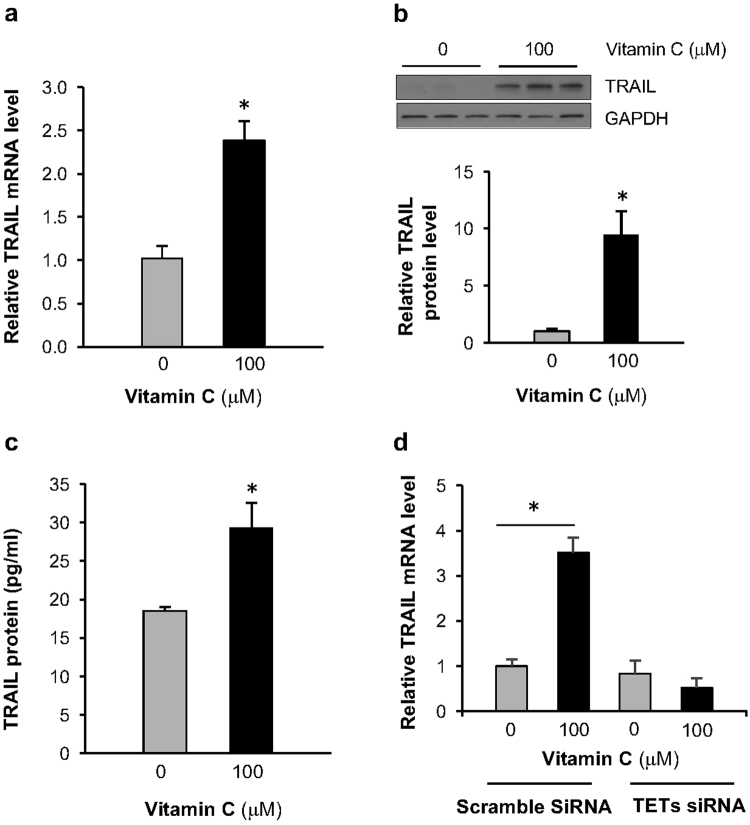
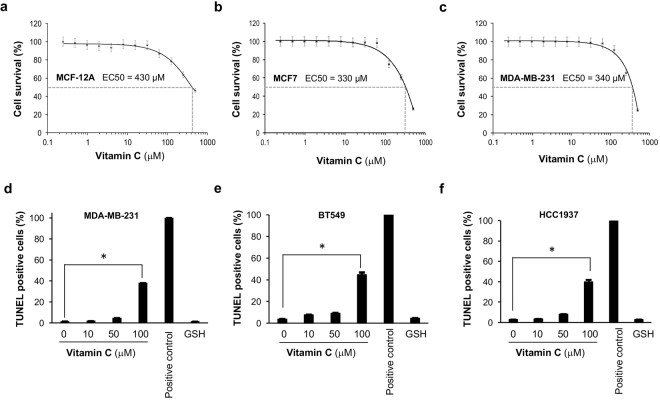
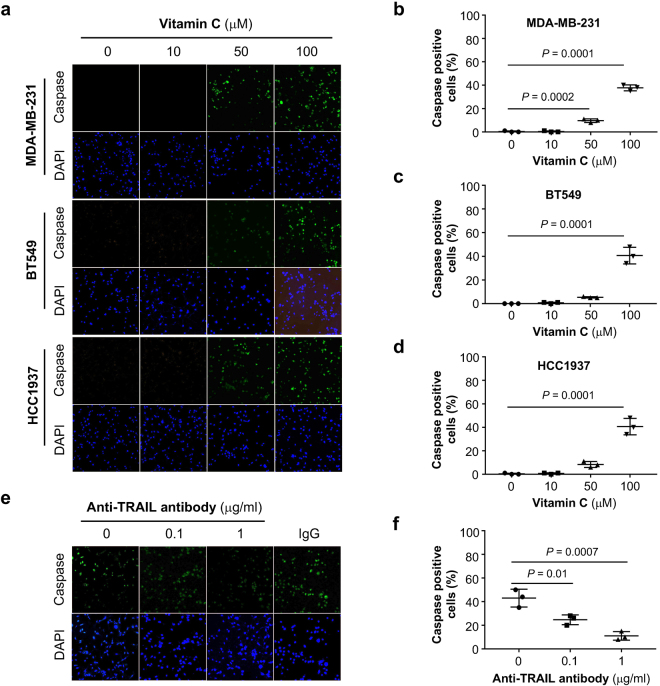
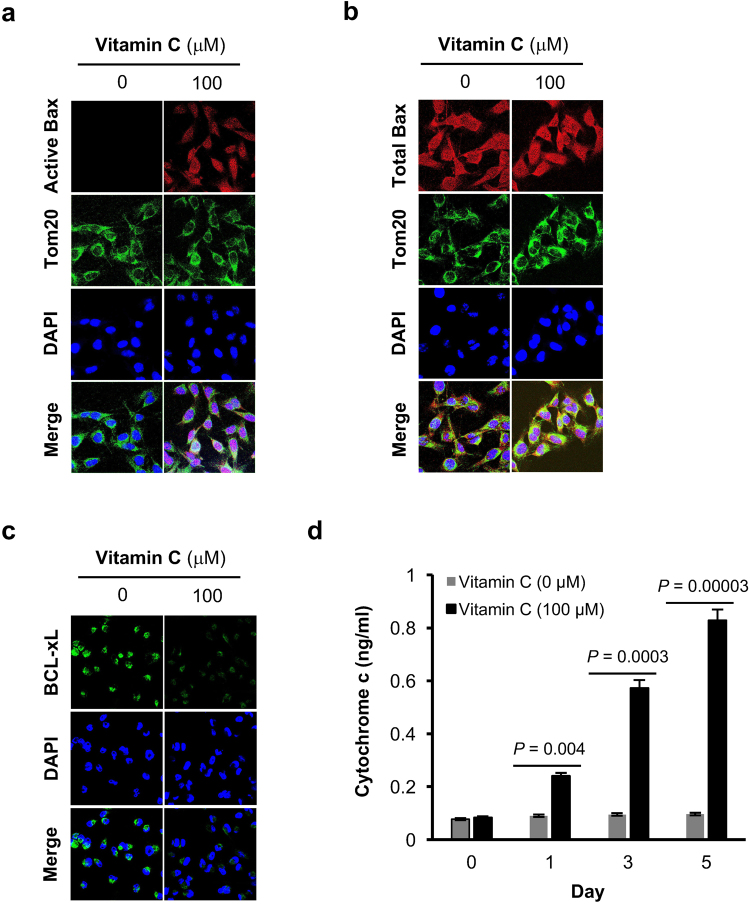
Genomic loss of 5-hydroxymethylcytosine (5hmC) accompanies malignant cellular transformation in breast cancer. Vitamin C serves as a cofactor for TET methylcytosine dioxygenases to increase 5hmC generation. Here we show that the transcription of SVCT2, a major vitamin C transporter, was decreased in human breast cancers (113 cases) compared to normal breast tissues from the same patients. A decreased SVCT2 expression was also observed in breast cancer cell lines. Treatment with vitamin C (100 μM) increased the 5hmC content in MDA-MB-231 breast cancer cells and markedly altered the transcriptome. The vitamin C treatment induced apoptosis in MDA-MB-231 cells, which was verified in two additional breast cancer cell lines. This pro-apoptotic effect of vitamin C appeared to be mediated by TRAIL, a known apoptosis inducer. Vitamin C upregulated TRAIL transcripts (2.3-fold increase) and increased TRAIL protein levels. The upregulation of TRAIL by vitamin C was largely abolished by siRNAs targeting TETs and anti-TRAIL antibody abrogated the induction of apoptosis. Furthermore, the apoptosis promoted by vitamin C was associated with Bax and caspases activation, Bcl-xL sequestration, and cytochrome c release. Taken together, these results suggest a potential role of physiological doses of vitamin C in breast cancer prevention and treatment.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
