

Review
doi: 10.1038/s41556-018-0067-6.
Epub 2018 Mar 28.
Deconstructing the pluripotency gene regulatory network
Affiliations
- PMID: 29593328
- PMCID: PMC6620196
- DOI: 10.1038/s41556-018-0067-6
Item in Clipboard
Review
Deconstructing the pluripotency gene regulatory network
Nat Cell Biol.
2018 Apr.
Abstract
Pluripotent stem cells can be isolated from embryos or derived by reprogramming. Pluripotency is stabilized by an interconnected network of pluripotency genes that cooperatively regulate gene expression. Here we describe the molecular principles of pluripotency gene function and highlight post-transcriptional controls, particularly those induced by RNA-binding proteins and alternative splicing, as an important regulatory layer of pluripotency. We also discuss heterogeneity in pluripotency regulation, alternative pluripotency states and future directions of pluripotent stem cell research.
Conflict of interest statement
Competing interests
The authors declare no competing interests.
Figures
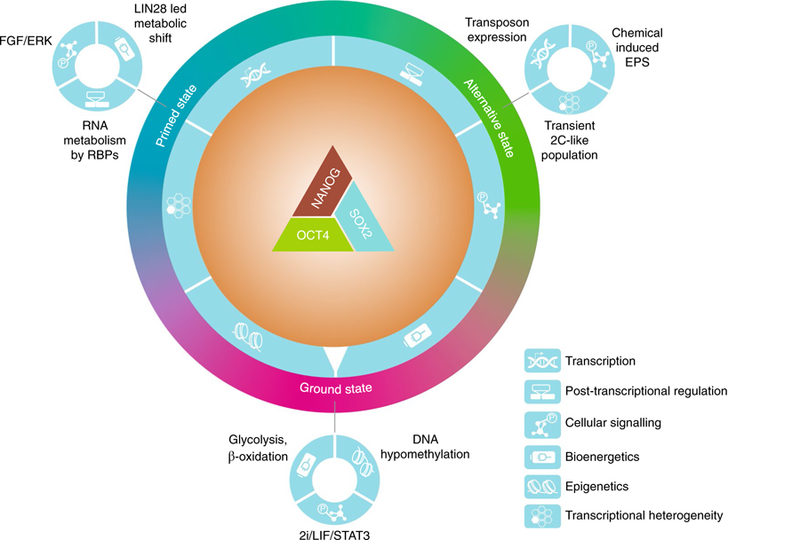
Pluripotency is stabilized by a triad of core transcription factors; namely OCT4, SOX2 and NANOG, which act cooperatively to regulate a larger and interconnected network of pluripotency genes. The PGRN crosstalks with multiple regulatory mechanisms, including transcription, post-transcriptional regulation, cellular signalling, bioenergetics, epigenetics and transcriptional heterogeneity (depicted with symbols on a dial outside of the core PGRN). For example, LIN28 is a PSC-associated RBP that mediates a metabolic shift from naïve to primed pluripotency by targeting mRNA translation, while the stability of LIN28 itself is controlled by fibroblast growth factor (FGF)–ERK signalling. The integration of all regulatory inputs ultimately ‘dials’ PSCs in specific pluripotent states, such as the ground state, primed state and alternative pluripotency states. The primed, ground and alternative states are depicted as a colour spectrum because evidence suggests that in vivo pluripotency exists as a dynamic continuum and that these states are interconvertible in vitro.
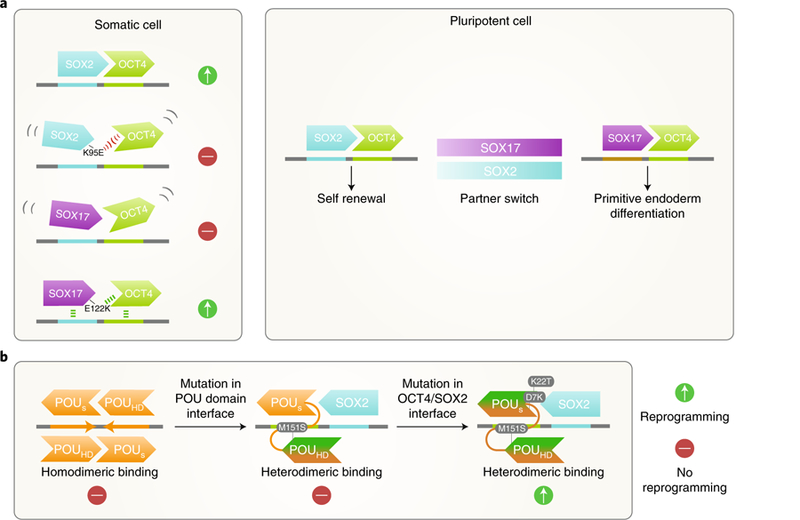
a, Key residues at the OCT4 interaction interface of SOX family transcription factors determine their ability to cooperatively bind with OCT4 and reprogram somatic cells to iPSCs. In PSCs, when the availability of SOX2 becomes limited, OCT4 can switch to partner with SOX17 and engage alternative heterodimeric motifs to promote differentiation. b, Other POU family members of OCT4, such as OCT6 (depicted in orange), prefer to homodimerize on the palindromic OCT–OCT motif. Mutagenesis of a key residue (M151S) in the interface between the POU-specific domain (POUs) and C-terminal POU homodomain (POUHD) of OCT6 redirects it to the heterodimeric OCT–SOX motif. Two additional mutations in the OCT4–SOX2 interaction interface in the POUs domain convert OCT6 into a reprogramming factor.
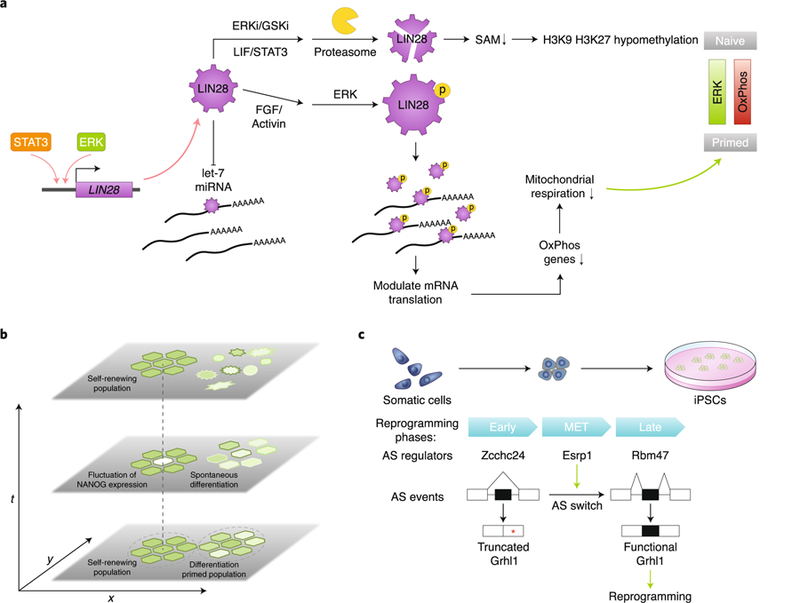
a, LIN28 levels in PSCs are regulated at both the transcriptional and post-transcriptional levels. It opposes let-7 microRNA biogenesis and also has let-7 independent functions. LIN28 can coordinate upstream signalling and post-transcriptional control to regulate the metabolic and epigenetic state of PSCs. b, Transcriptional heterogeneity in PSCs manifests as transcriptional fluctuation over time, as seen in the case of NANOG. This is possibly caused by transcriptional noise and autoregulatory loops between pluripotency transcription factors. Heterogeneity in PSCs can also originate from subpopulation structure. For example, self-renewing PSCs can coexist with subpopulations of cells with reduced pluripotency gene expression and a higher propensity for differentiation over time. c, Multiphasic alternative splicing (AS) program changes during somatic cell reprogramming. Many AS regulators contribute to phase-specific AS changes during reprogramming, including Zcchc24 for the early phase, Esrp1 for the MET phase and Rbm47 for the late phase. Esrp1 promotes inclusion of an alternatively spliced exon in Grhl1, which produces a functional protein that facilitates MET rather than a truncated form. The full-length Grhl1 enhances reprogramming efficiency. Red asterisk, stop codon; OxPhos, oxidative phosphorylation; SAM, S-adenosyl-methionine.
References
-
- Huang Y, Osorno R, Tsakiridis A & Wilson V In vivo differentiation potential of epiblast stem cells revealed by chimeric embryo formation. Cell Rep 2, 1571–1578 (2012). - PubMed
-
- Rosenthal MD, Wishnow RM & Sato GH In vitro growth and differetiation of clonal populations of multipotential mouse clls derived from a transplantable testicular teratocarcinoma. J. Natl. Cancer I 44, 1001–1014 (1970). - PubMed
-
- Evans MJ & Kaufman MH Establishment in culture of pluripotential cells from mouse embryos. Nature 292, 154–156 (1981). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
