

Profiles of miRNA Isoforms and tRNA Fragments in Prostate Cancer
- PMID: 29593348
- PMCID: PMC5871839
- DOI: 10.1038/s41598-018-22488-2
Profiles of miRNA Isoforms and tRNA Fragments in Prostate Cancer
Abstract
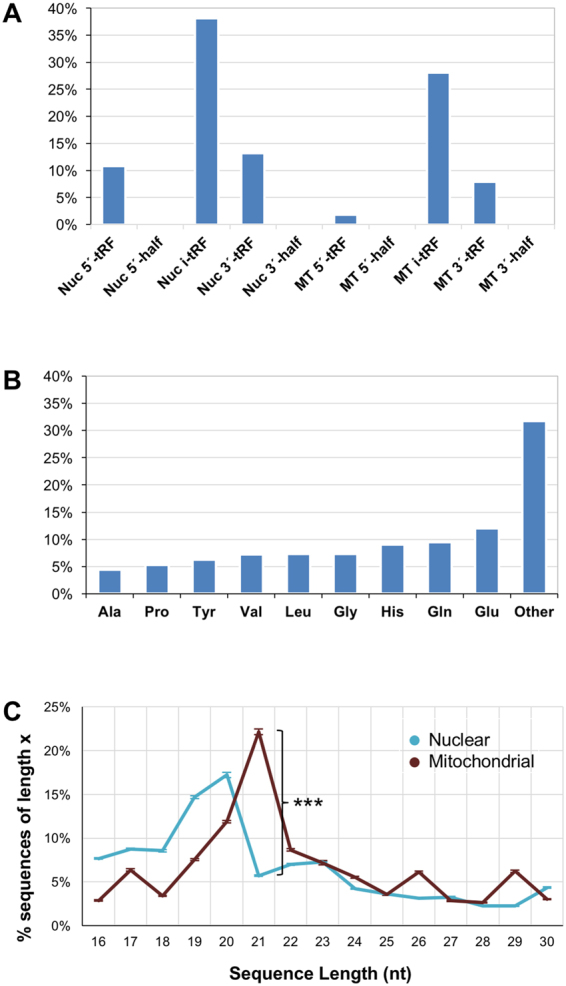
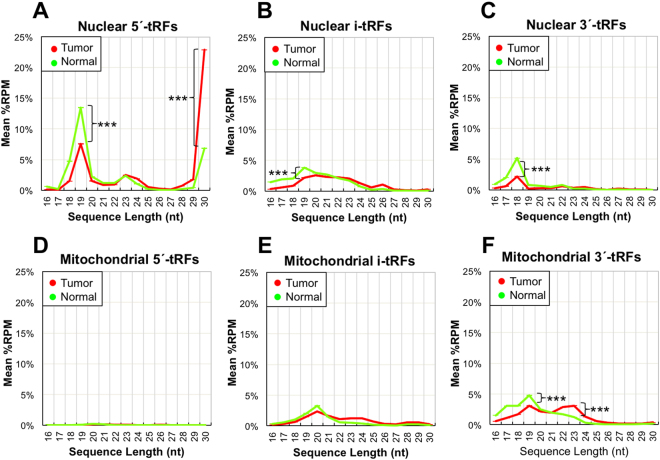
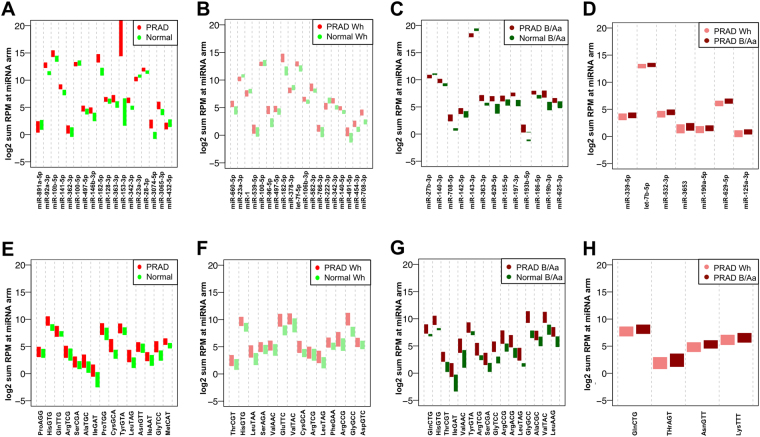
MicroRNA (miRNA) isoforms ("isomiRs") and tRNA-derived fragments ("tRFs") are powerful regulatory non-coding RNAs (ncRNAs). In human tissues, both types of molecules are abundant, with expression patterns that depend on a person's race, sex and population origin. Here, we present our analyses of the Prostate Cancer (PRAD) datasets of The Cancer Genome Atlas (TCGA) from the standpoint of isomiRs and tRFs. This study represents the first simultaneous examination of isomiRs and tRFs in a large cohort of PRAD patients. We find that isomiRs and tRFs have extensive correlations with messenger RNAs (mRNAs). These correlations are disrupted in PRAD, which suggests disruptions of the regulatory network in the disease state. Notably, we find that the profiles of isomiRs and tRFs differ in patients belonging to different races. We hope that the presented findings can lay the groundwork for future research efforts aimed at elucidating the functional roles of the numerous and distinct members of these two categories of ncRNAs that are present in PRAD.
Conflict of interest statement
The authors declare no competing interests.
Figures




References
-
- Kayani, M., Kayani, M. A., Malik, F. A. & Faryal, R. Role of miRNAs in breast cancer. Asian Pac J Cancer Prev12 (2011). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
