

Enhanced Expression of the Key Mitosis Regulator Cyclin B1 Is Mediated by PDZ-Binding Kinase in Islets of Pregnant Mice
- PMID: 29594255
- PMCID: PMC5841183
- DOI: 10.1210/js.2017-00338
Enhanced Expression of the Key Mitosis Regulator Cyclin B1 Is Mediated by PDZ-Binding Kinase in Islets of Pregnant Mice
Abstract
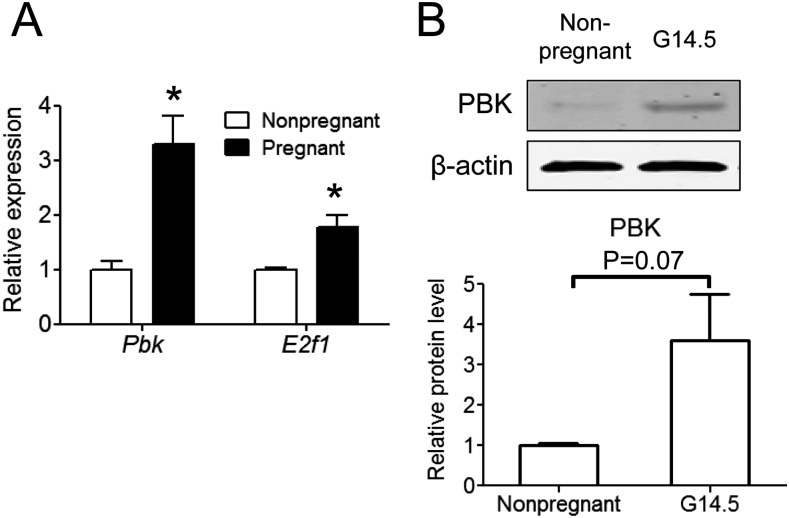
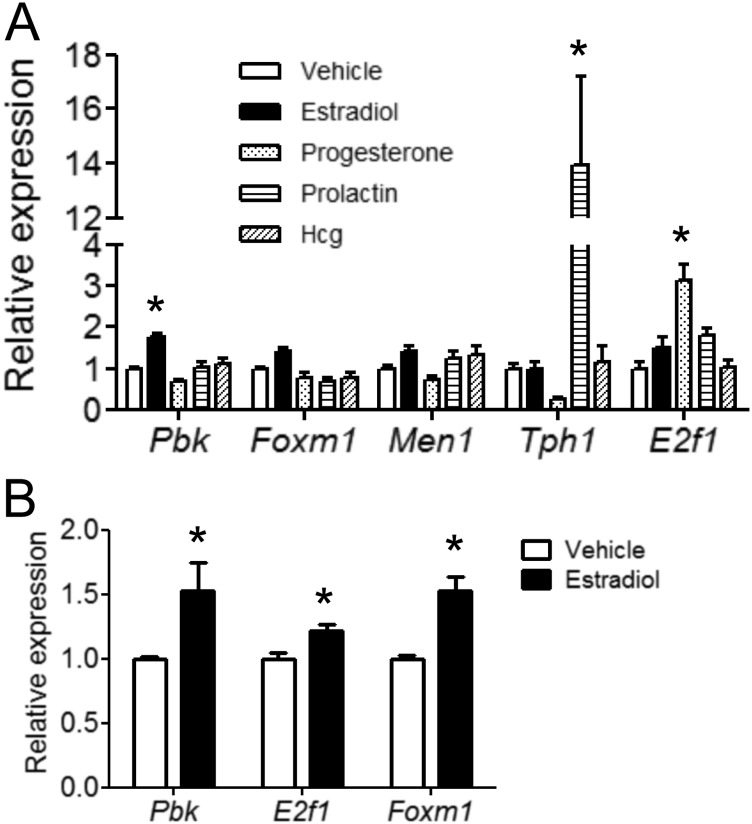
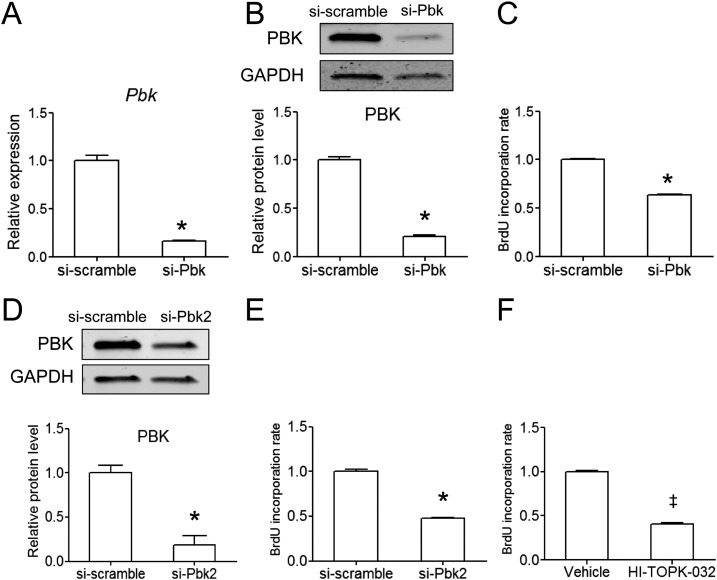
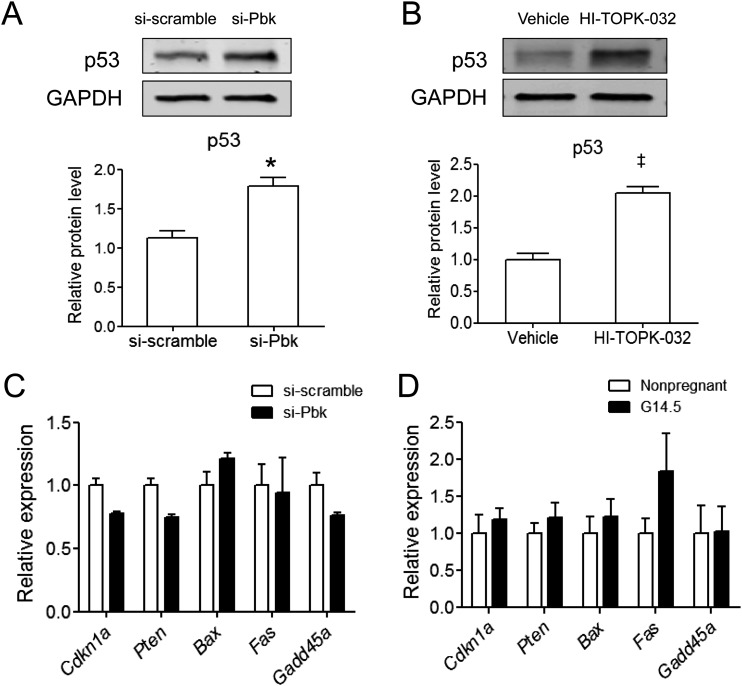
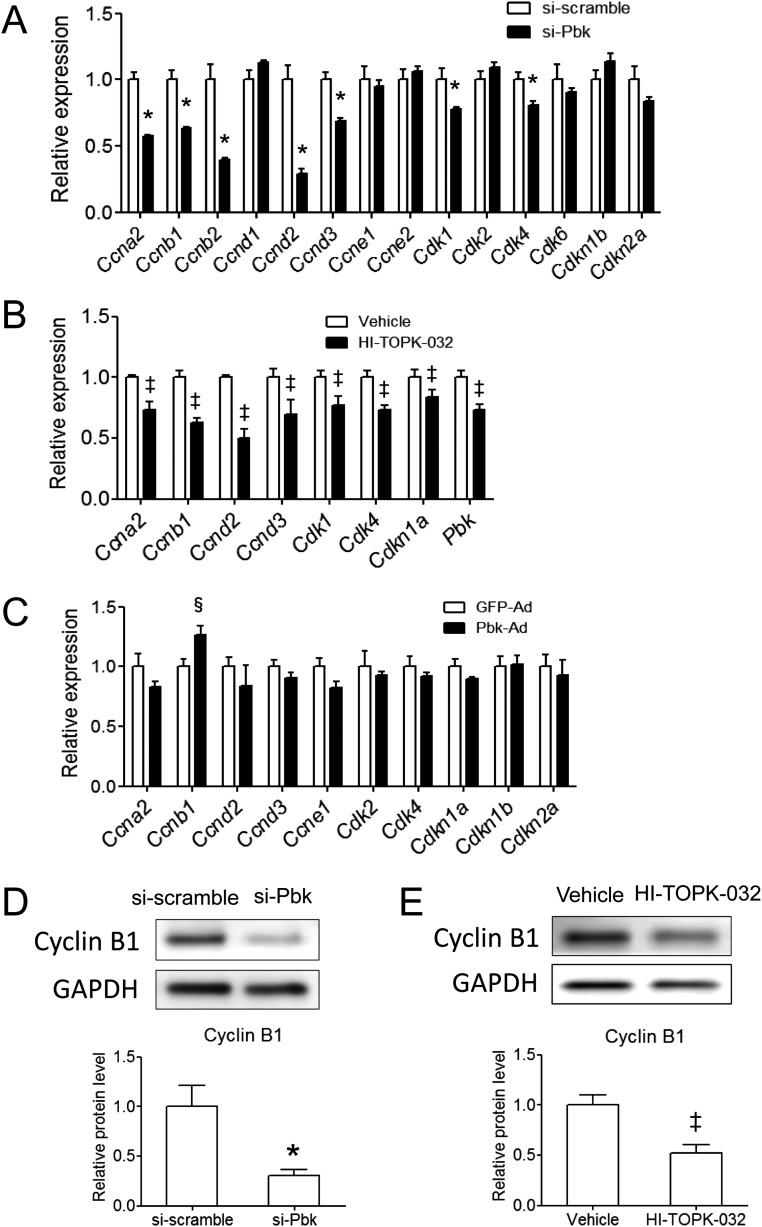
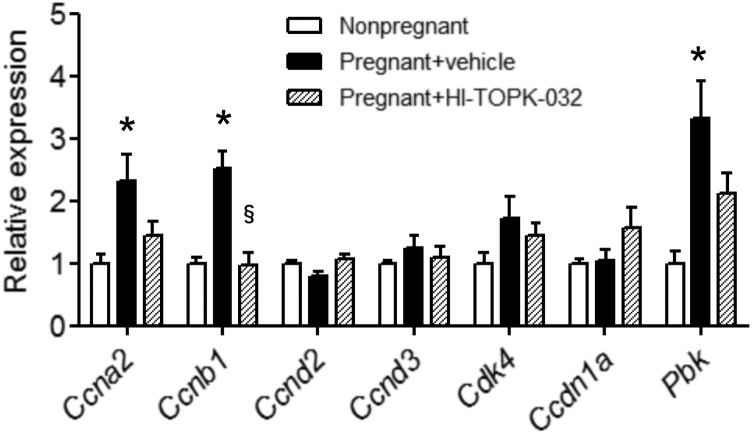
The proliferation of pancreatic β cells is enhanced to enable an increase in β-cell mass and to compensate for insulin resistance during pregnancy. To elucidate the mechanisms involved, we previously investigated islets from pregnant and nonpregnant mice by gene expression profiling and found that the expression of postsynaptic density-95/Discs large/zonula occludens-1 (PDZ)-binding kinase (Pbk), a member of the mitogen-activated protein kinase kinase family, is increased in pregnant mouse islets compared with control mouse islets. Among the pregnancy hormones, treatment with estradiol upregulated Pbk expression. Inhibition of Pbk expression using a small interfering RNA for Pbk reduced bromodeoxyuridine incorporation in mouse insulinoma 6 cells, which was accompanied by a decreased expression of Ccnb1, a regulatory gene involved in mitosis. Ccnb1 expression was augmented in mouse islets during pregnancy. The forced expression of Pbk using an adenovirus system in isolated mouse islets increased Ccnb1 expression, and the Pbk inhibitor HI-TOPK-032 suppressed Ccnb1 expression in islets isolated from pregnant mice. Our results suggest that Pbk contributes to the expansion of islets during pregnancy and that Ccnb1 may assist Pbk in its role in β-cell proliferation.
Keywords: Pbk; cyclin B1; estradiol; islet; p53.
Conflict of interest statement
T.O. has received research funding from AstraZeneca, Eli Lilly and Company, Sanofi-Aventis, and Mitsubishi Tanabe Pharma. The remaining authors have nothing to disclose.
Figures
References
-
- Huang C, Snider F, Cross JC. Prolactin receptor is required for normal glucose homeostasis and modulation of beta-cell mass during pregnancy. Endocrinology. 2009;150(4):1618–1626. - PubMed
-
- Hughes E, Huang C. Participation of Akt, menin, and p21 in pregnancy-induced beta-cell proliferation. Endocrinology. 2011;152(3):847–855. - PubMed
-
- Karnik SK, Chen H, McLean GW, Heit JJ, Gu X, Zhang AY, Fontaine M, Yen MH, Kim SK. Menin controls growth of pancreatic beta-cells in pregnant mice and promotes gestational diabetes mellitus. Science. 2007;318(5851):806–809. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous