

Structure of the nucleotide exchange factor eIF2B reveals mechanism of memory-enhancing molecule
- PMID: 29599213
- PMCID: PMC6120582
- DOI: 10.1126/science.aaq0939
Structure of the nucleotide exchange factor eIF2B reveals mechanism of memory-enhancing molecule
Abstract
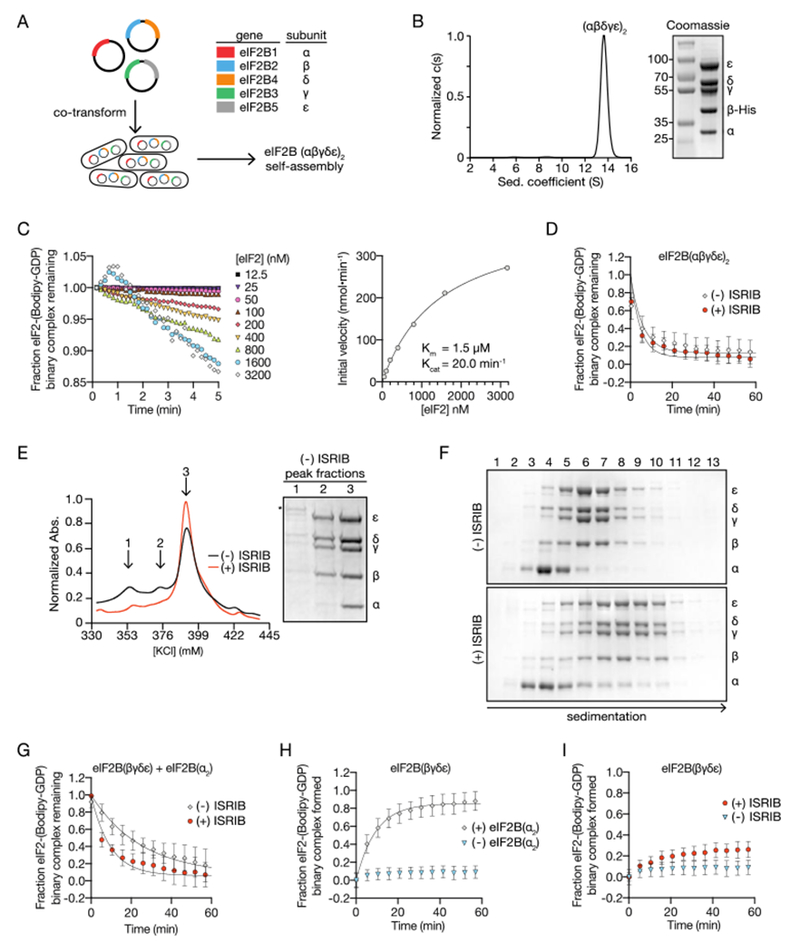
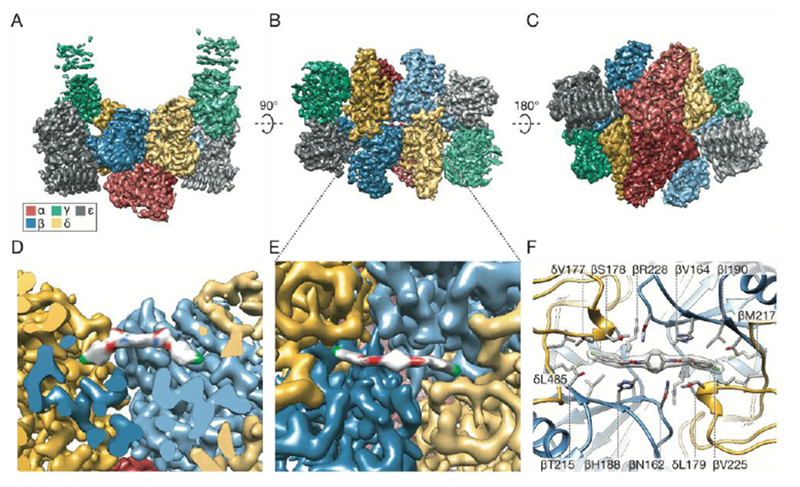
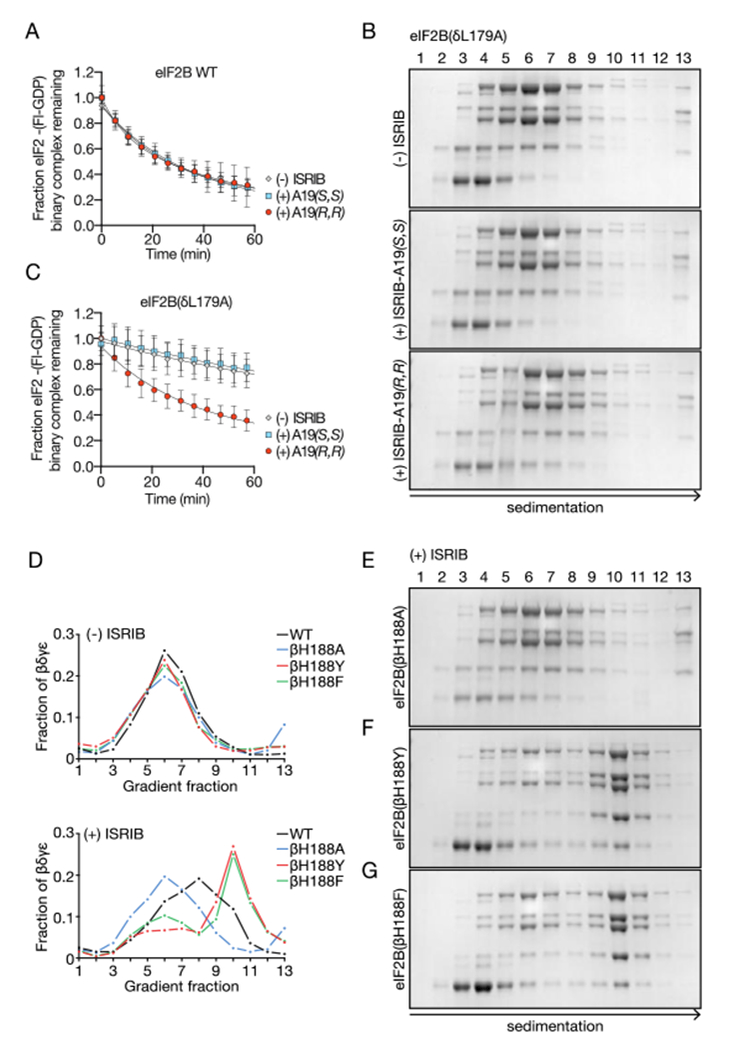
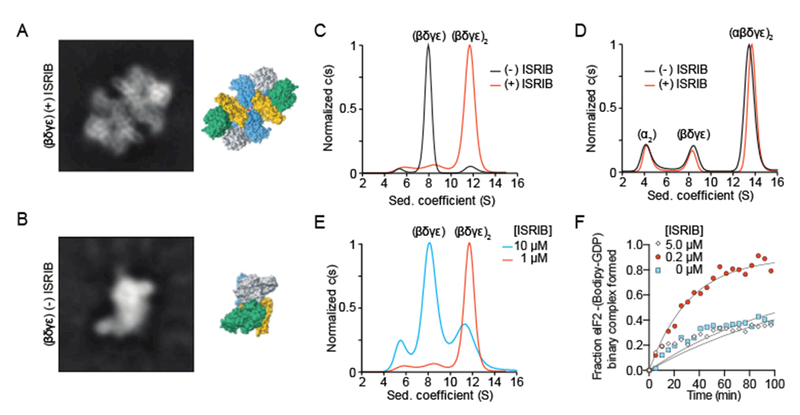
Regulation by the integrated stress response (ISR) converges on the phosphorylation of translation initiation factor eIF2 in response to a variety of stresses. Phosphorylation converts eIF2 from a substrate to a competitive inhibitor of its dedicated guanine nucleotide exchange factor, eIF2B, thereby inhibiting translation. ISRIB, a drug-like eIF2B activator, reverses the effects of eIF2 phosphorylation, and in rodents it enhances cognition and corrects cognitive deficits after brain injury. To determine its mechanism of action, we solved an atomic-resolution structure of ISRIB bound in a deep cleft within decameric human eIF2B by cryo-electron microscopy. Formation of fully active, decameric eIF2B holoenzyme depended on the assembly of two identical tetrameric subcomplexes, and ISRIB promoted this step by cross-bridging a central symmetry interface. Thus, regulation of eIF2B assembly emerges as a rheostat for eIF2B activity that tunes translation during the ISR and that can be further modulated by ISRIB.
Copyright © 2018 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Figures
Similar articles
-
Structural insights into ISRIB, a memory-enhancing inhibitor of the integrated stress response.FEBS J. 2020 Jan;287(2):239-245. doi: 10.1111/febs.15073. Epub 2019 Nov 7. FEBS J. 2020. PMID: 31550413 Review.
-
Pharmacological dimerization and activation of the exchange factor eIF2B antagonizes the integrated stress response.Elife. 2015 Apr 15;4:e07314. doi: 10.7554/eLife.07314. Elife. 2015. PMID: 25875391 Free PMC article.
-
Binding of ISRIB reveals a regulatory site in the nucleotide exchange factor eIF2B.Science. 2018 Mar 30;359(6383):1533-1536. doi: 10.1126/science.aar5129. Science. 2018. PMID: 29599245 Free PMC article.
-
ISRIB Blunts the Integrated Stress Response by Allosterically Antagonising the Inhibitory Effect of Phosphorylated eIF2 on eIF2B.Mol Cell. 2021 Jan 7;81(1):88-103.e6. doi: 10.1016/j.molcel.2020.10.031. Epub 2020 Nov 20. Mol Cell. 2021. PMID: 33220178 Free PMC article.
-
Protection of eIF2B from inhibitory phosphorylated eIF2: A viral strategy to maintain mRNA translation during the PKR-triggered integrated stress response.J Biol Chem. 2023 Nov;299(11):105287. doi: 10.1016/j.jbc.2023.105287. Epub 2023 Sep 22. J Biol Chem. 2023. PMID: 37742919 Free PMC article. Review.
Cited by
-
Integrated Stress Response Inhibitor Reverses Sex-Dependent Behavioral and Cell-Specific Deficits after Mild Repetitive Head Trauma.J Neurotrauma. 2020 Jun 1;37(11):1370-1380. doi: 10.1089/neu.2019.6827. Epub 2020 Feb 11. J Neurotrauma. 2020. PMID: 31884883 Free PMC article.
-
The Human Cytomegalovirus Nonstructural Glycoprotein UL148 Reorganizes the Endoplasmic Reticulum.mBio. 2019 Dec 10;10(6):e02110-19. doi: 10.1128/mBio.02110-19. mBio. 2019. PMID: 31822584 Free PMC article.
-
Regulation and function of elF2B in neurological and metabolic disorders.Biosci Rep. 2022 Jun 30;42(6):BSR20211699. doi: 10.1042/BSR20211699. Biosci Rep. 2022. PMID: 35579296 Free PMC article. Review.
-
Formation and persistence of polyglutamine aggregates in mistranslating cells.Nucleic Acids Res. 2021 Nov 18;49(20):11883-11899. doi: 10.1093/nar/gkab898. Nucleic Acids Res. 2021. PMID: 34718744 Free PMC article.
-
Targeting Protein Synthesis in Colorectal Cancer.Cancers (Basel). 2020 May 21;12(5):1298. doi: 10.3390/cancers12051298. Cancers (Basel). 2020. PMID: 32455578 Free PMC article. Review.
References
-
- Munn DH et al., GCN2 kinase in T cells mediates proliferative arrest and anergy induction in response to indoleamine 2,3-dioxygenase. Immunity. 22, 633–642 (2005). - PubMed
-
- Wortham NC, Martinez M, Gordiyenko Y, Robinson CV, Proud CG, Analysis of the subunit organization of the eIF2B complex reveals new insights into its structure and regulation. The FASEB Journal. 28, 2225–2237 (2014). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
