

Erythrocytic ferroportin reduces intracellular iron accumulation, hemolysis, and malaria risk
- PMID: 29599243
- PMCID: PMC8349187
- DOI: 10.1126/science.aal2022
Erythrocytic ferroportin reduces intracellular iron accumulation, hemolysis, and malaria risk
Abstract
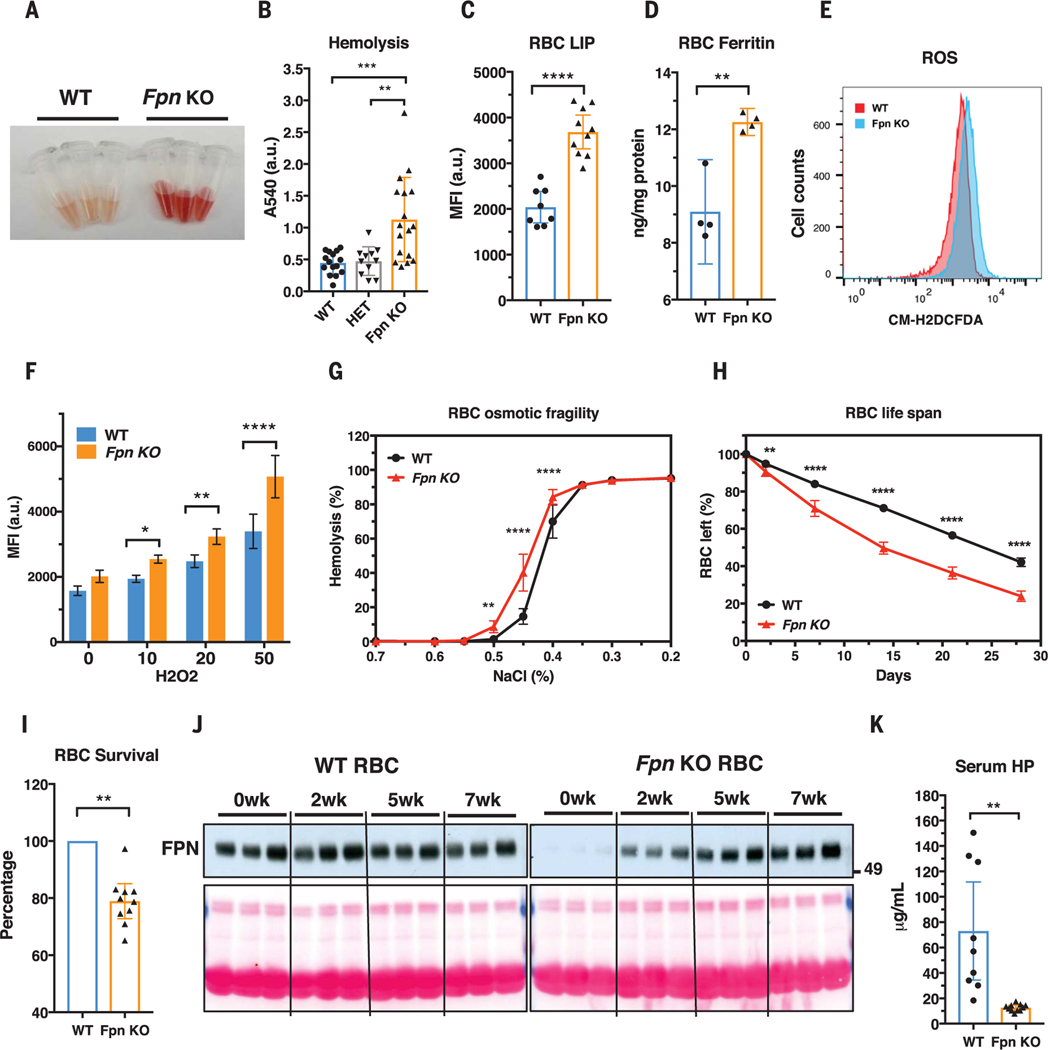
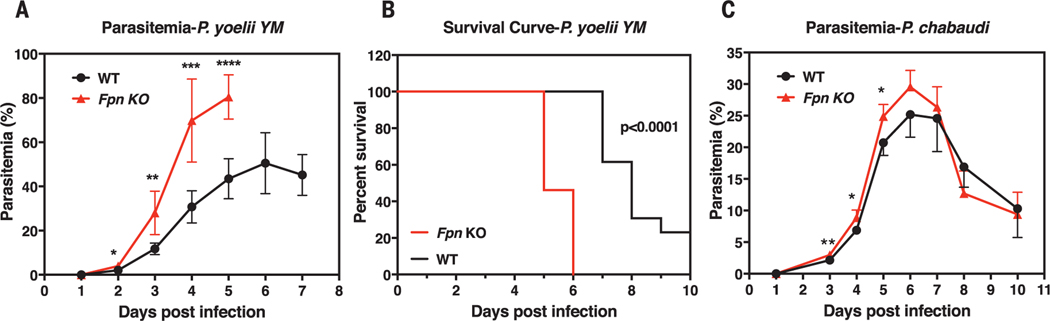
Malaria parasites invade red blood cells (RBCs), consume copious amounts of hemoglobin, and severely disrupt iron regulation in humans. Anemia often accompanies malaria disease; however, iron supplementation therapy inexplicably exacerbates malarial infections. Here we found that the iron exporter ferroportin (FPN) was highly abundant in RBCs, and iron supplementation suppressed its activity. Conditional deletion of the Fpn gene in erythroid cells resulted in accumulation of excess intracellular iron, cellular damage, hemolysis, and increased fatality in malaria-infected mice. In humans, a prevalent FPN mutation, Q248H (glutamine to histidine at position 248), prevented hepcidin-induced degradation of FPN and protected against severe malaria disease. FPN Q248H appears to have been positively selected in African populations in response to the impact of malaria disease. Thus, FPN protects RBCs against oxidative stress and malaria infection.
Copyright © 2018 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
