

Helraiser intermediates provide insight into the mechanism of eukaryotic replicative transposition
- PMID: 29599430
- PMCID: PMC5876387
- DOI: 10.1038/s41467-018-03688-w
Helraiser intermediates provide insight into the mechanism of eukaryotic replicative transposition
Abstract
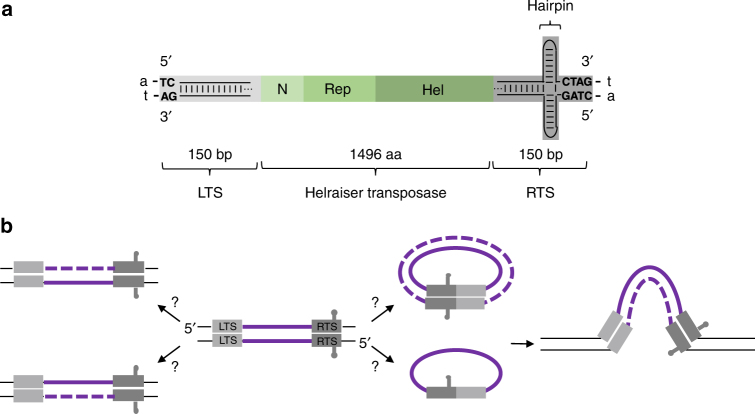
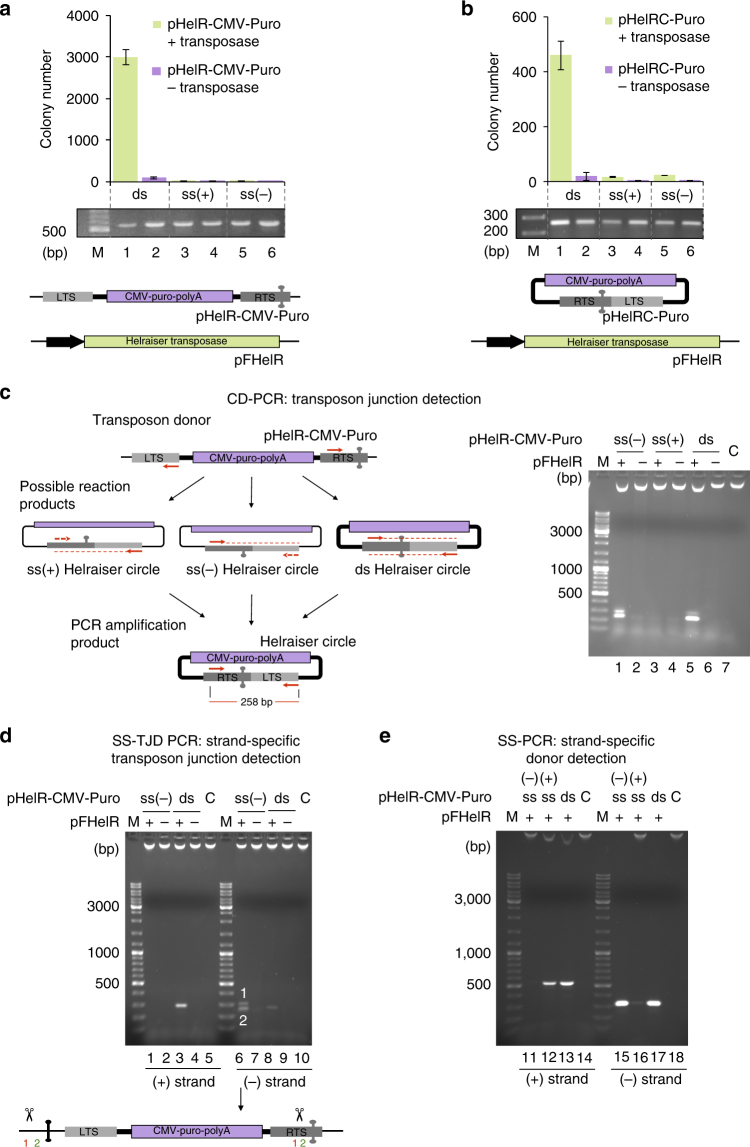
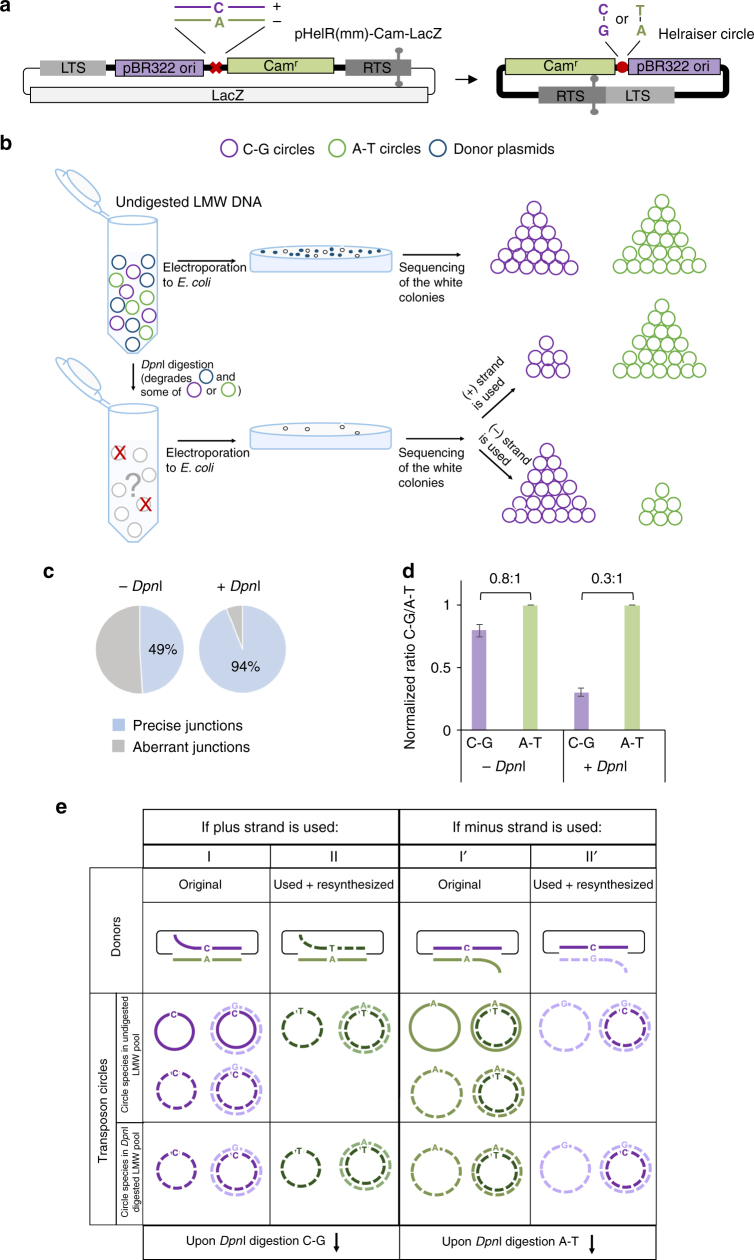
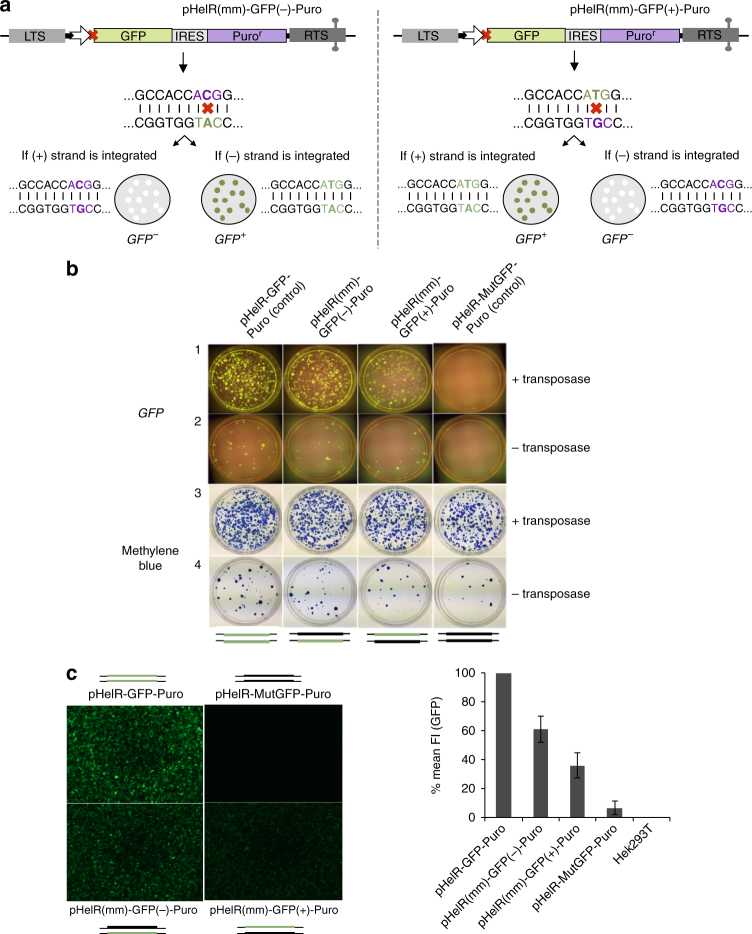
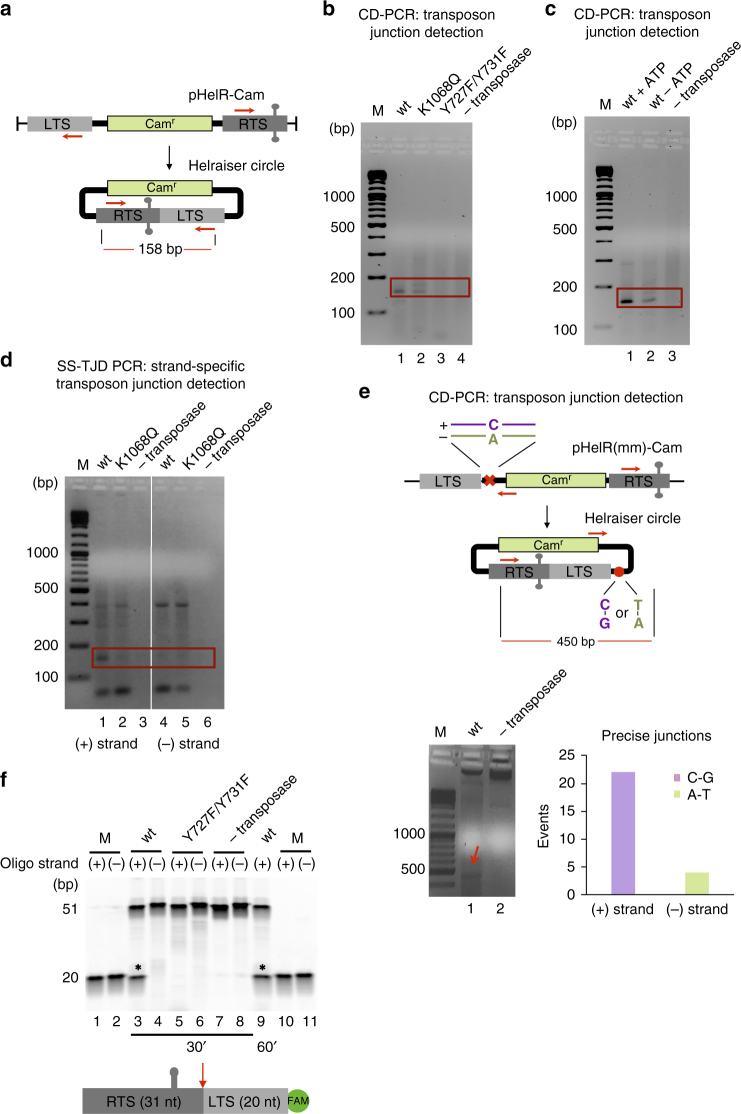
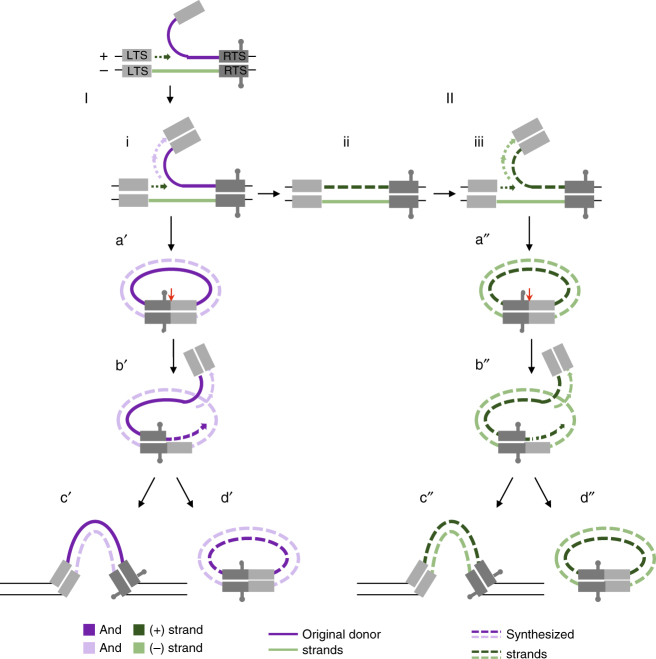
Helitrons are eukaryotic DNA transposons that have profoundly affected genome variability via capture and mobilization of host genomic sequences. Defining their mode of action is therefore important for understanding how genome landscapes evolve. Sequence similarities with certain prokaryotic mobile elements suggest a "rolling circle" mode of transposition, involving only a single transposon strand. Using the reconstituted Helraiser transposon to study Helitron transposition in cells and in vitro, we show that the donor site must be double-stranded and that single-stranded donors will not suffice. Nevertheless, replication and integration assays demonstrate the use of only one of the transposon donor strands. Furthermore, repeated reuse of Helraiser donor sites occurs following DNA synthesis. In cells, circular double-stranded intermediates that serve as transposon donors are generated and replicated by Helraiser transposase. Cell-free experiments demonstrate strand-specific cleavage and strand transfer, supporting observations made in cells.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
A Helitron transposon reconstructed from bats reveals a novel mechanism of genome shuffling in eukaryotes.Nat Commun. 2016 Mar 2;7:10716. doi: 10.1038/ncomms10716. Nat Commun. 2016. PMID: 26931494 Free PMC article.
-
Transposition of hAT elements links transposable elements and V(D)J recombination.Nature. 2004 Dec 23;432(7020):995-1001. doi: 10.1038/nature03157. Nature. 2004. PMID: 15616554
-
The large bat Helitron DNA transposase forms a compact monomeric assembly that buries and protects its covalently bound 5'-transposon end.Mol Cell. 2021 Oct 21;81(20):4271-4286.e4. doi: 10.1016/j.molcel.2021.07.028. Epub 2021 Aug 16. Mol Cell. 2021. PMID: 34403695 Free PMC article.
-
[Transposition as a way of existence: phage Mu].Genetika. 2003 May;39(5):637-56. Genetika. 2003. PMID: 12838611 Review. Russian.
-
Sleeping Beauty Transposition.Microbiol Spectr. 2015 Apr;3(2):MDNA3-0042-2014. doi: 10.1128/microbiolspec.MDNA3-0042-2014. Microbiol Spectr. 2015. PMID: 26104705 Review.
Cited by
-
Diversification of the Caenorhabditis heat shock response by Helitron transposable elements.Elife. 2019 Dec 11;8:e51139. doi: 10.7554/eLife.51139. Elife. 2019. PMID: 31825311 Free PMC article.
-
Zinc-finger BED domains drive the formation of the active Hermes transpososome by asymmetric DNA binding.Nat Commun. 2023 Jul 25;14(1):4470. doi: 10.1038/s41467-023-40210-3. Nat Commun. 2023. PMID: 37491363 Free PMC article.
-
Coexistence vs collapse in transposon populations.ArXiv [Preprint]. 2025 May 19:arXiv:2411.11010v2. ArXiv. 2025. Update in: Phys Rev E. 2025 May;111(5-1):054414. doi: 10.1103/PhysRevE.111.054414. PMID: 40470469 Free PMC article. Updated. Preprint.
-
RepeatModeler2 for automated genomic discovery of transposable element families.Proc Natl Acad Sci U S A. 2020 Apr 28;117(17):9451-9457. doi: 10.1073/pnas.1921046117. Epub 2020 Apr 16. Proc Natl Acad Sci U S A. 2020. PMID: 32300014 Free PMC article.
-
Exploring the Remote Ties between Helitron Transposases and Other Rolling-Circle Replication Proteins.Int J Mol Sci. 2018 Oct 9;19(10):3079. doi: 10.3390/ijms19103079. Int J Mol Sci. 2018. PMID: 30304800 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
