

RodZ modulates geometric localization of the bacterial actin MreB to regulate cell shape
- PMID: 29599448
- PMCID: PMC5876373
- DOI: 10.1038/s41467-018-03633-x
RodZ modulates geometric localization of the bacterial actin MreB to regulate cell shape
Abstract
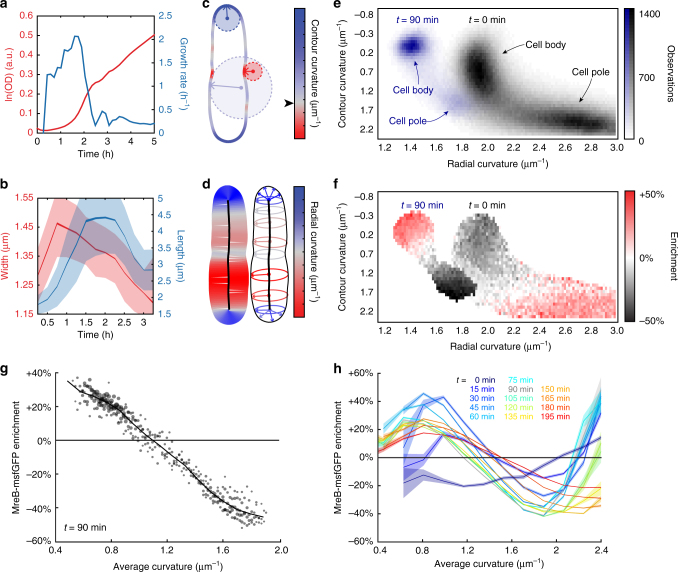
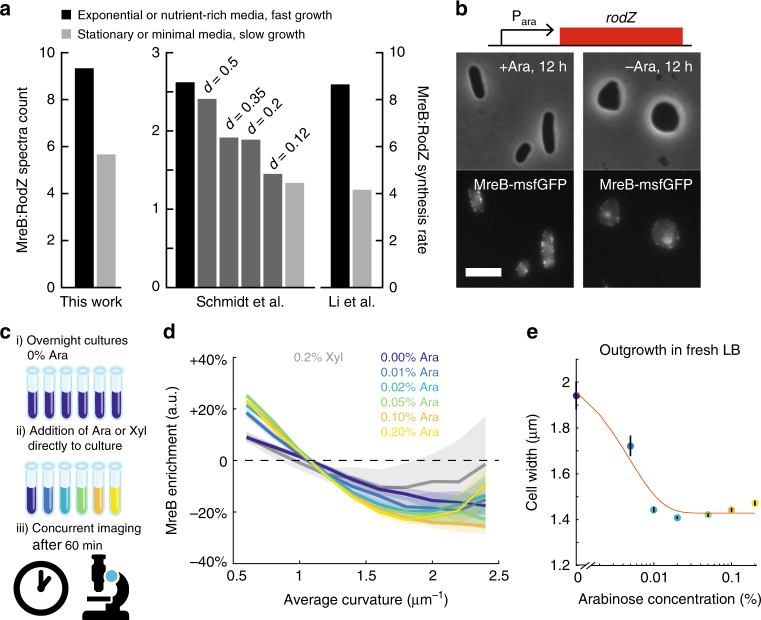
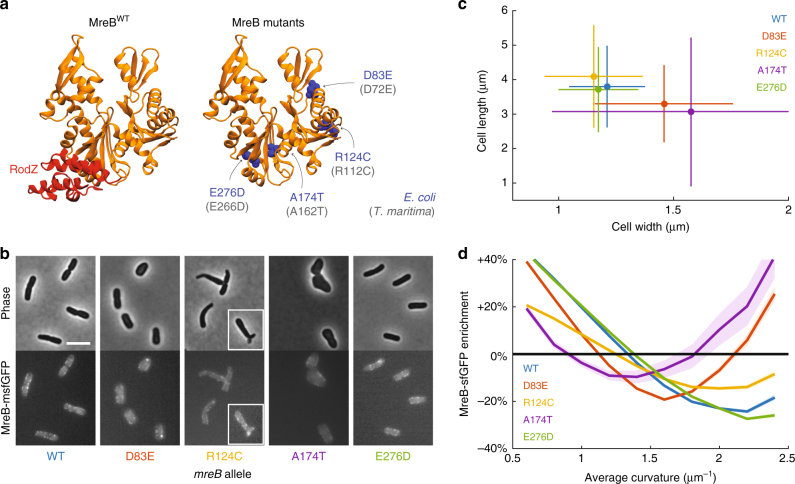
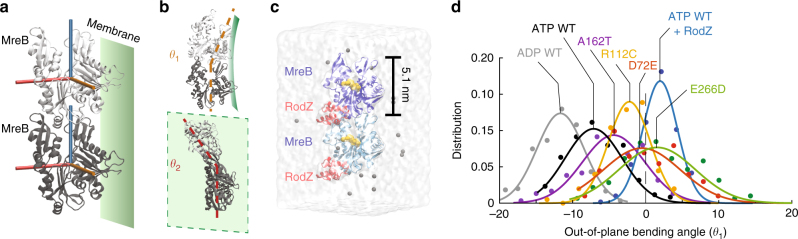
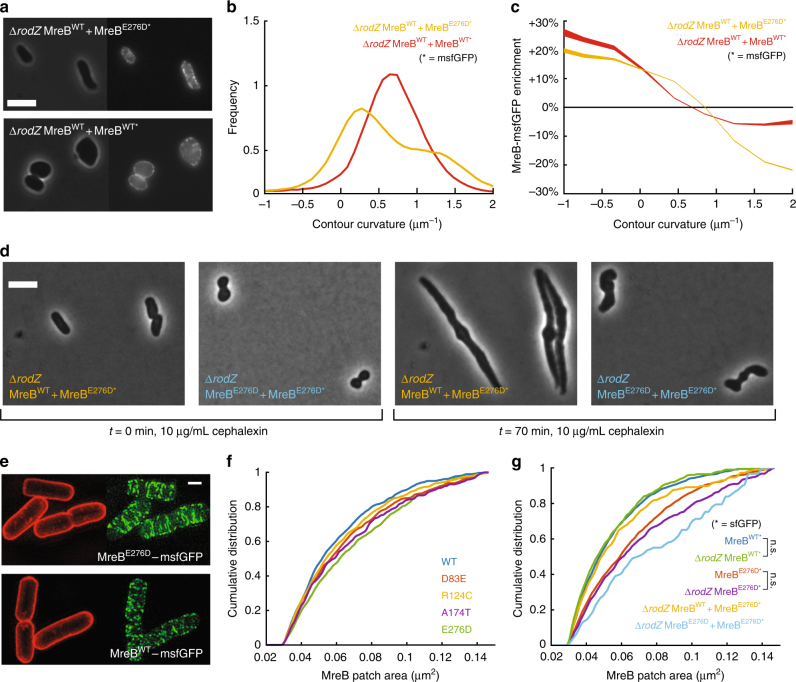
In the rod-shaped bacterium Escherichia coli, the actin-like protein MreB localizes in a curvature-dependent manner and spatially coordinates cell-wall insertion to maintain cell shape, although the molecular mechanism by which cell width is regulated remains unknown. Here we demonstrate that the membrane protein RodZ regulates the biophysical properties of MreB and alters the spatial organization of E. coli cell-wall growth. The relative expression levels of MreB and RodZ change in a manner commensurate with variations in growth rate and cell width, and RodZ systematically alters the curvature-based localization of MreB and cell width in a concentration-dependent manner. We identify MreB mutants that alter the bending properties of MreB filaments in molecular dynamics simulations similar to RodZ binding, and show that these mutants rescue rod-like shape in the absence of RodZ alone or in combination with wild-type MreB. Thus, E. coli can control its shape and dimensions by differentially regulating RodZ and MreB to alter the patterning of cell-wall insertion, highlighting the rich regulatory landscape of cytoskeletal molecular biophysics.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
How to Build a Bacterial Cell: MreB as the Foreman of E. coli Construction.Cell. 2018 Mar 8;172(6):1294-1305. doi: 10.1016/j.cell.2018.02.050. Cell. 2018. PMID: 29522748 Free PMC article. Review.
-
RodZ links MreB to cell wall synthesis to mediate MreB rotation and robust morphogenesis.Proc Natl Acad Sci U S A. 2015 Oct 6;112(40):12510-5. doi: 10.1073/pnas.1509610112. Epub 2015 Sep 22. Proc Natl Acad Sci U S A. 2015. PMID: 26396257 Free PMC article.
-
Chlamydial MreB Directs Cell Division and Peptidoglycan Synthesis in Escherichia coli in the Absence of FtsZ Activity.mBio. 2020 Feb 18;11(1):e03222-19. doi: 10.1128/mBio.03222-19. mBio. 2020. PMID: 32071268 Free PMC article.
-
The periplasmic disordered domain of RodZ promotes its self-interaction in Escherichia coli.Genes Cells. 2018 Apr;23(4):307-317. doi: 10.1111/gtc.12572. Epub 2018 Feb 26. Genes Cells. 2018. PMID: 29480545
-
[Regulation of determination of bacterial shape].Nihon Saikingaku Zasshi. 2014;69(4):557-64. doi: 10.3412/jsb.69.557. Nihon Saikingaku Zasshi. 2014. PMID: 25447981 Review. Japanese.
Cited by
-
Surface-to-volume scaling and aspect ratio preservation in rod-shaped bacteria.Elife. 2019 Aug 28;8:e47033. doi: 10.7554/eLife.47033. Elife. 2019. PMID: 31456563 Free PMC article.
-
How to Build a Bacterial Cell: MreB as the Foreman of E. coli Construction.Cell. 2018 Mar 8;172(6):1294-1305. doi: 10.1016/j.cell.2018.02.050. Cell. 2018. PMID: 29522748 Free PMC article. Review.
-
Environmental and Physiological Factors Affecting High-Throughput Measurements of Bacterial Growth.mBio. 2020 Oct 20;11(5):e01378-20. doi: 10.1128/mBio.01378-20. mBio. 2020. PMID: 33082255 Free PMC article.
-
A specialized MreB-dependent cell wall biosynthetic complex mediates the formation of stalk-specific peptidoglycan in Caulobacter crescentus.PLoS Genet. 2019 Feb 1;15(2):e1007897. doi: 10.1371/journal.pgen.1007897. eCollection 2019 Feb. PLoS Genet. 2019. PMID: 30707707 Free PMC article.
-
MreB: unraveling the molecular mechanisms of bacterial shape, division, and environmental adaptation.Cell Commun Signal. 2025 Aug 22;23(1):377. doi: 10.1186/s12964-025-02373-y. Cell Commun Signal. 2025. PMID: 40847400 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
