

Transcript Profiling Identifies NAC-Domain Genes Involved in Regulating Wall Ingrowth Deposition in Phloem Parenchyma Transfer Cells of Arabidopsis thaliana
- PMID: 29599795
- PMCID: PMC5862824
- DOI: 10.3389/fpls.2018.00341
Transcript Profiling Identifies NAC-Domain Genes Involved in Regulating Wall Ingrowth Deposition in Phloem Parenchyma Transfer Cells of Arabidopsis thaliana
Abstract
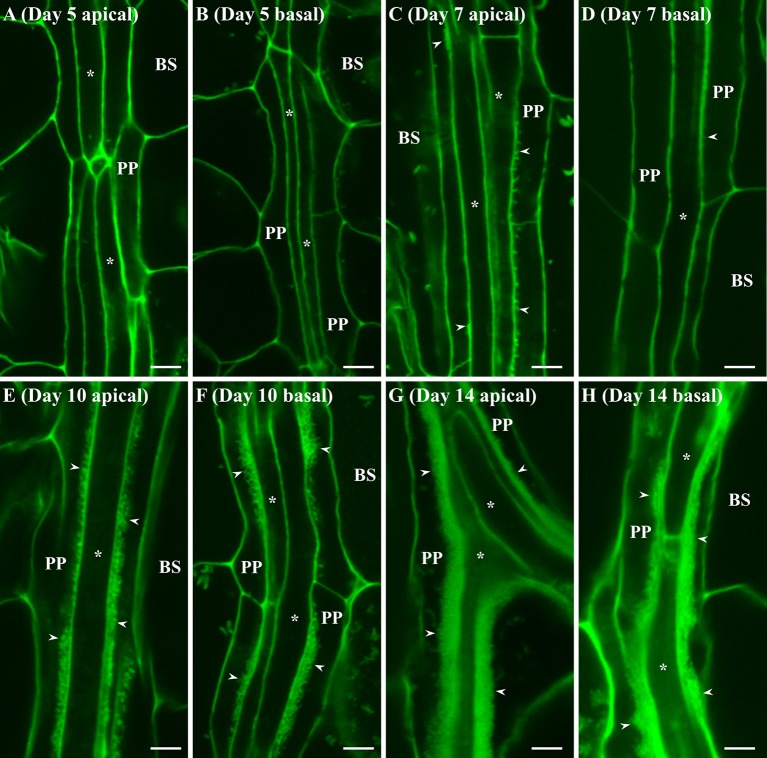
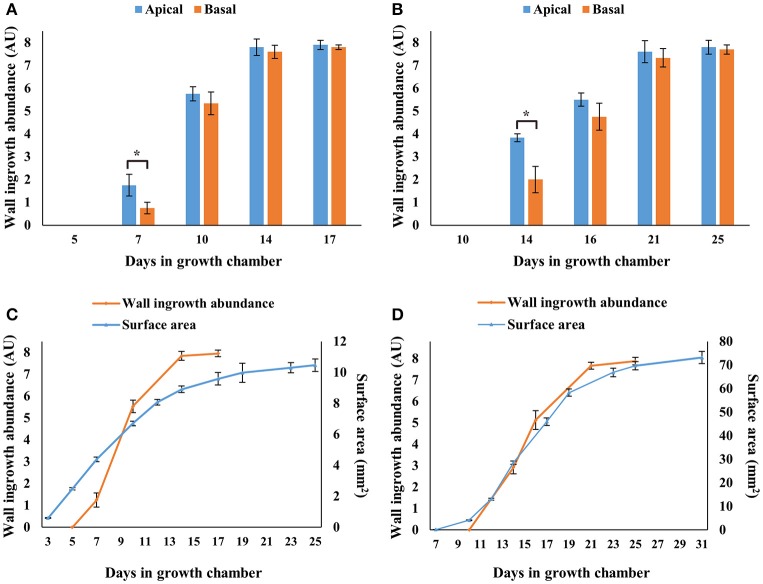
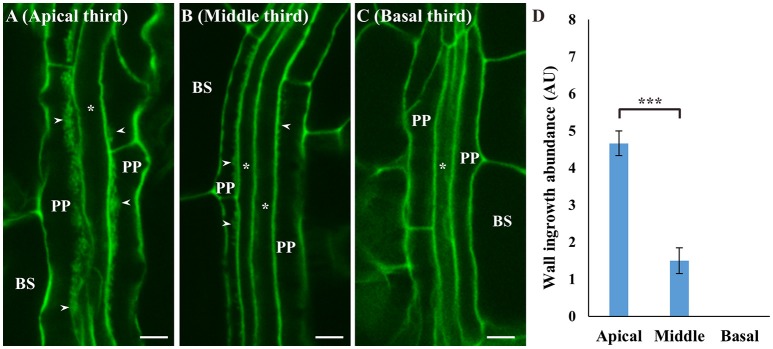
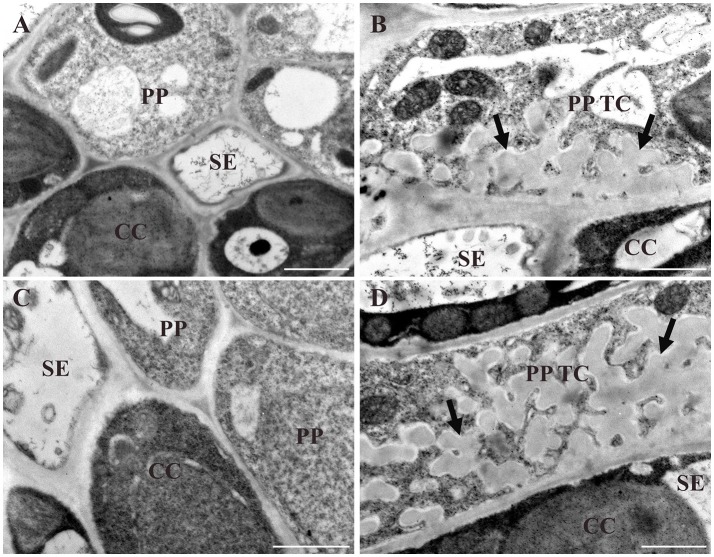
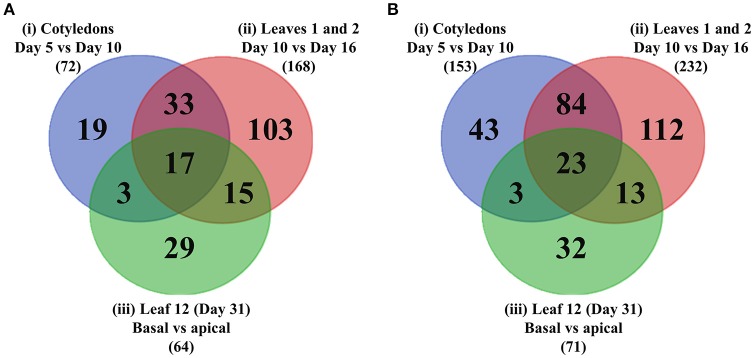
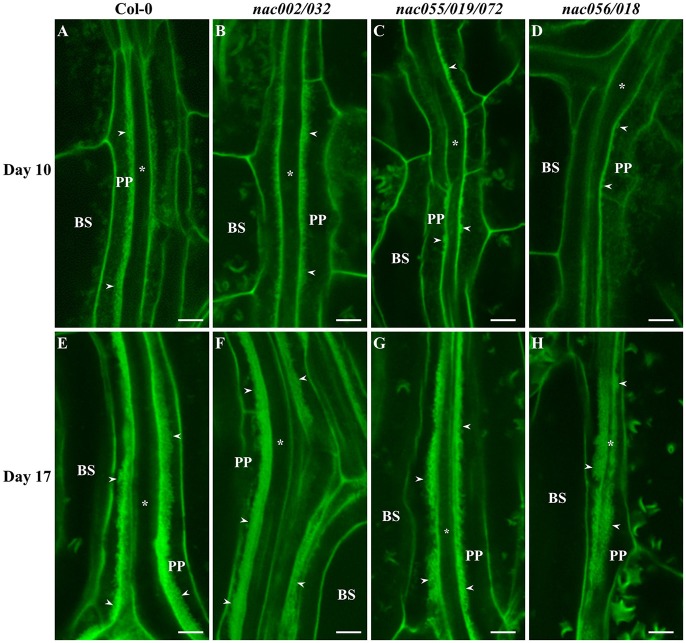
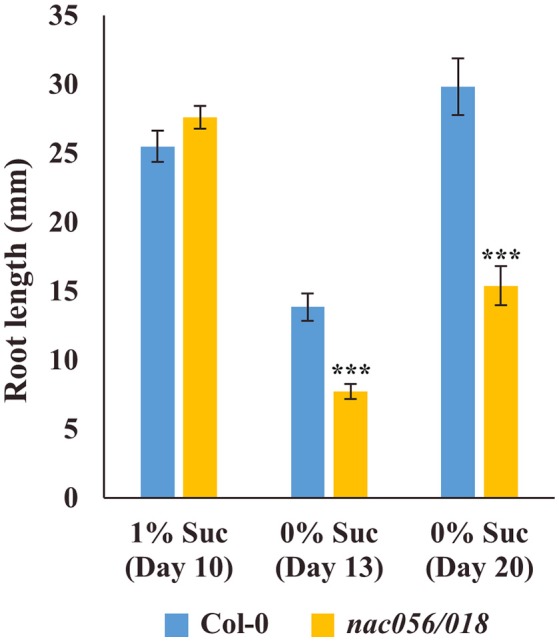
Transfer cells (TCs) play important roles in facilitating enhanced rates of nutrient transport at key apoplasmic/symplasmic junctions along the nutrient acquisition and transport pathways in plants. TCs achieve this capacity by developing elaborate wall ingrowth networks which serve to increase plasma membrane surface area thus increasing the cell's surface area-to-volume ratio to achieve increased flux of nutrients across the plasma membrane. Phloem parenchyma (PP) cells of Arabidopsis leaf veins trans-differentiate to become PP TCs which likely function in a two-step phloem loading mechanism by facilitating unloading of photoassimilates into the apoplasm for subsequent energy-dependent uptake into the sieve element/companion cell (SE/CC) complex. We are using PP TCs in Arabidopsis as a genetic model to identify transcription factors involved in coordinating deposition of the wall ingrowth network. Confocal imaging of pseudo-Schiff propidium iodide-stained tissue revealed different profiles of temporal development of wall ingrowth deposition across maturing cotyledons and juvenile leaves, and a basipetal gradient of deposition across mature adult leaves. RNA-Seq analysis was undertaken to identify differentially expressed genes common to these three different profiles of wall ingrowth deposition. This analysis identified 68 transcription factors up-regulated two-fold or more in at least two of the three experimental comparisons, with six of these transcription factors belonging to Clade III of the NAC-domain family. Phenotypic analysis of these NAC genes using insertional mutants revealed significant reductions in levels of wall ingrowth deposition, particularly in a double mutant of NAC056 and NAC018, as well as compromised sucrose-dependent root growth, indicating impaired capacity for phloem loading. Collectively, these results support the proposition that Clade III members of the NAC-domain family in Arabidopsis play important roles in regulating wall ingrowth deposition in PP TCs.
Keywords: Arabidopsis thaliana; RNA-Seq; phloem parenchyma; transcription factors; transfer cells; wall ingrowths.
Figures



Similar articles
-
High-resolution confocal imaging of wall ingrowth deposition in plant transfer cells: Semi-quantitative analysis of phloem parenchyma transfer cell development in leaf minor veins of Arabidopsis.BMC Plant Biol. 2015 Apr 23;15:109. doi: 10.1186/s12870-015-0483-8. BMC Plant Biol. 2015. PMID: 25899055 Free PMC article.
-
Review: More than sweet: New insights into the biology of phloem parenchyma transfer cells in Arabidopsis.Plant Sci. 2021 Sep;310:110990. doi: 10.1016/j.plantsci.2021.110990. Epub 2021 Jul 8. Plant Sci. 2021. PMID: 34315604 Review.
-
Heteroblastic Development of Transfer Cells Is Controlled by the microRNA miR156/SPL Module.Plant Physiol. 2017 Mar;173(3):1676-1691. doi: 10.1104/pp.16.01741. Epub 2017 Jan 12. Plant Physiol. 2017. PMID: 28082719 Free PMC article.
-
Phloem parenchyma transfer cells in Arabidopsis - an experimental system to identify transcriptional regulators of wall ingrowth formation.Front Plant Sci. 2013 Apr 24;4:102. doi: 10.3389/fpls.2013.00102. eCollection 2013. Front Plant Sci. 2013. PMID: 23630536 Free PMC article.
-
Symplasmic transport and phloem loading in gymnosperm leaves.Protoplasma. 2011 Jan;248(1):181-90. doi: 10.1007/s00709-010-0239-0. Epub 2010 Nov 24. Protoplasma. 2011. PMID: 21107620 Free PMC article. Review.
Cited by
-
DNA methylation and its effects on gene expression during primary to secondary growth in poplar stems.BMC Genomics. 2020 Jul 20;21(1):498. doi: 10.1186/s12864-020-06902-6. BMC Genomics. 2020. PMID: 32689934 Free PMC article.
-
Distinct identities of leaf phloem cells revealed by single cell transcriptomics.Plant Cell. 2021 May 5;33(3):511-530. doi: 10.1093/plcell/koaa060. Plant Cell. 2021. PMID: 33955487 Free PMC article.
-
Lateral Transport of Organic and Inorganic Solutes.Plants (Basel). 2019 Jan 15;8(1):20. doi: 10.3390/plants8010020. Plants (Basel). 2019. PMID: 30650538 Free PMC article. Review.
-
Developmental Biology and Induction of Phi Thickenings by Abiotic Stress in Roots of the Brassicaceae.Plants (Basel). 2018 Jun 19;7(2):47. doi: 10.3390/plants7020047. Plants (Basel). 2018. PMID: 29921823 Free PMC article.
-
Differential Gene Expression in Contrasting Common Bean Cultivars for Drought Tolerance during an Extended Dry Period.Genes (Basel). 2024 Jul 17;15(7):935. doi: 10.3390/genes15070935. Genes (Basel). 2024. PMID: 39062714 Free PMC article.
References
-
- Amiard V., Mueh K. E., Demmig-Adams B., Ebbert V., Turgeon R., Adams W. W., III. (2005). Anatomical and photosynthetic acclimation to the light environment in species with differing mechanisms of phloem loading. Proc. Natl. Acad. Sci. U.S.A. 102, 12968–12973. 10.1073/pnas.0503784102 - DOI - PMC - PubMed
-
- Andriunas F. A., Zhang H. M., Weber H., McCurdy D. W., Offler C. E., Patrick J. W. (2011). Glucose and ethylene signalling pathways converge to regulate trans-differentiation of epidermal transfer cells in Vicia narbonensis cotyledons. Plant J. 68, 987–998. 10.1111/j.1365-313X.2011.04749.x - DOI - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous