

DNA Repair: The Search for Homology
- PMID: 29603285
- PMCID: PMC6238635
- DOI: 10.1002/bies.201700229
DNA Repair: The Search for Homology
Abstract
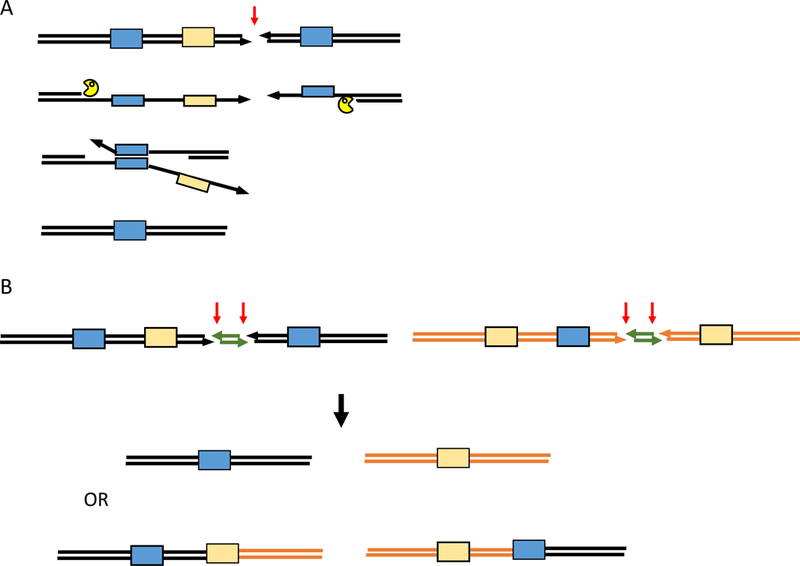
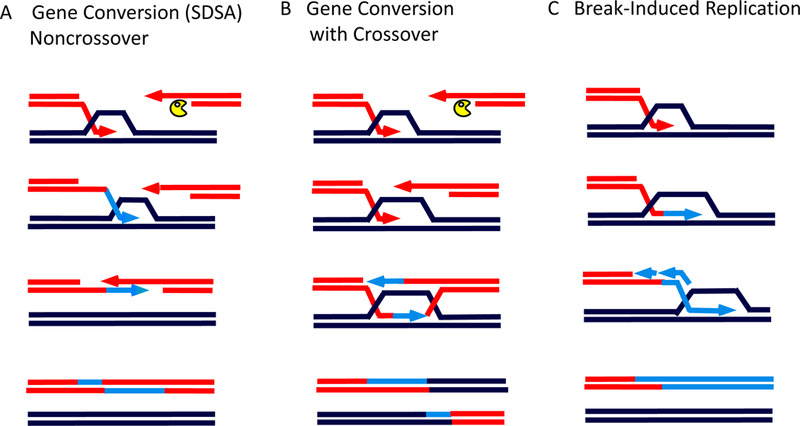
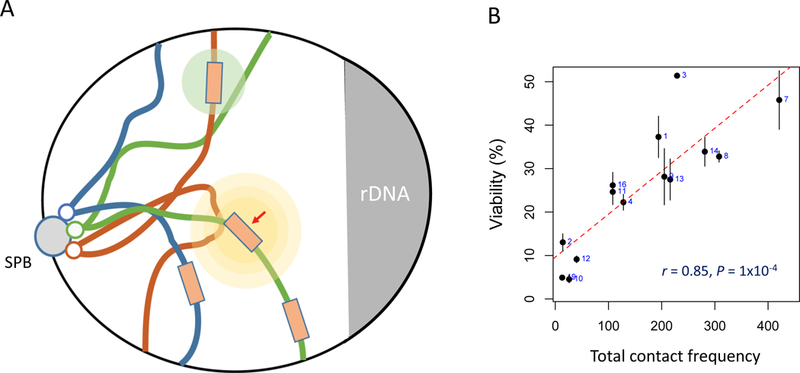
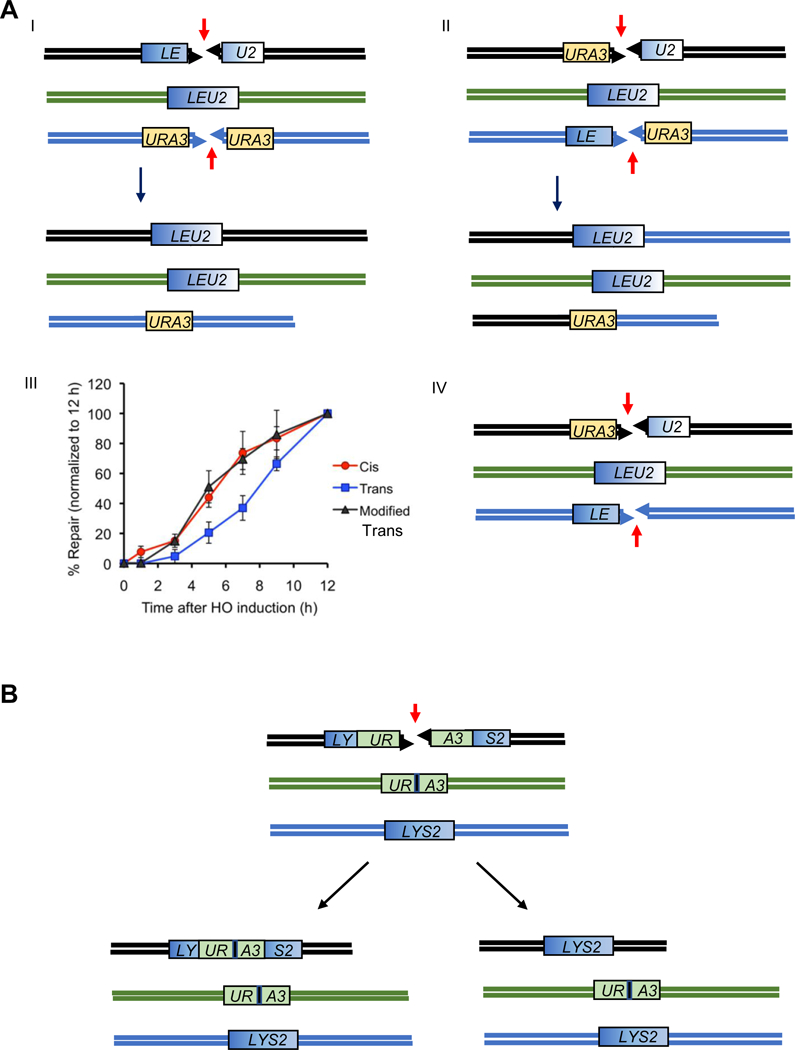
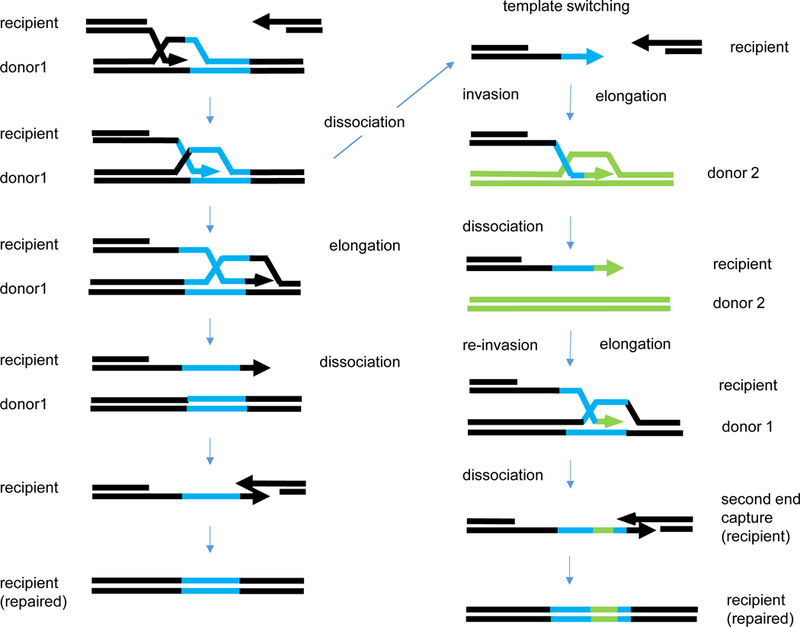
The repair of chromosomal double-strand breaks (DSBs) by homologous recombination is essential to maintain genome integrity. The key step in DSB repair is the RecA/Rad51-mediated process to match sequences at the broken end to homologous donor sequences that can be used as a template to repair the lesion. Here, in reviewing research about DSB repair, I consider the many factors that appear to play important roles in the successful search for homology by several homologous recombination mechanisms. See also the video abstract here: https://youtu.be/vm7-X5uIzS8.
Keywords: DNA strand invasion; Rad51/RecA; double-strand break repair; homologous recombination; search for homology.
© 2018 WILEY Periodicals, Inc.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
