

Regulation of C. elegans L4 cuticle collagen genes by the heterochronic protein LIN-29
- PMID: 29604168
- PMCID: PMC8672679
- DOI: 10.1002/dvg.23106
Regulation of C. elegans L4 cuticle collagen genes by the heterochronic protein LIN-29
Abstract
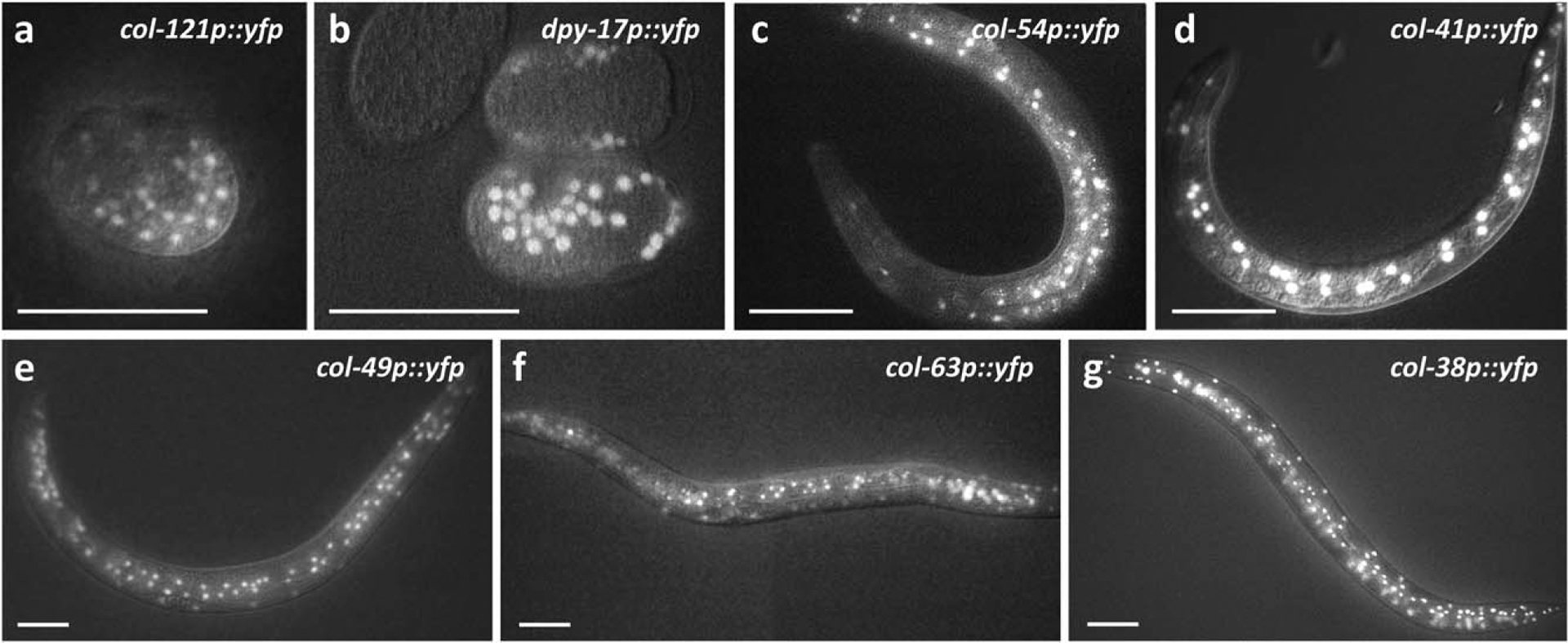
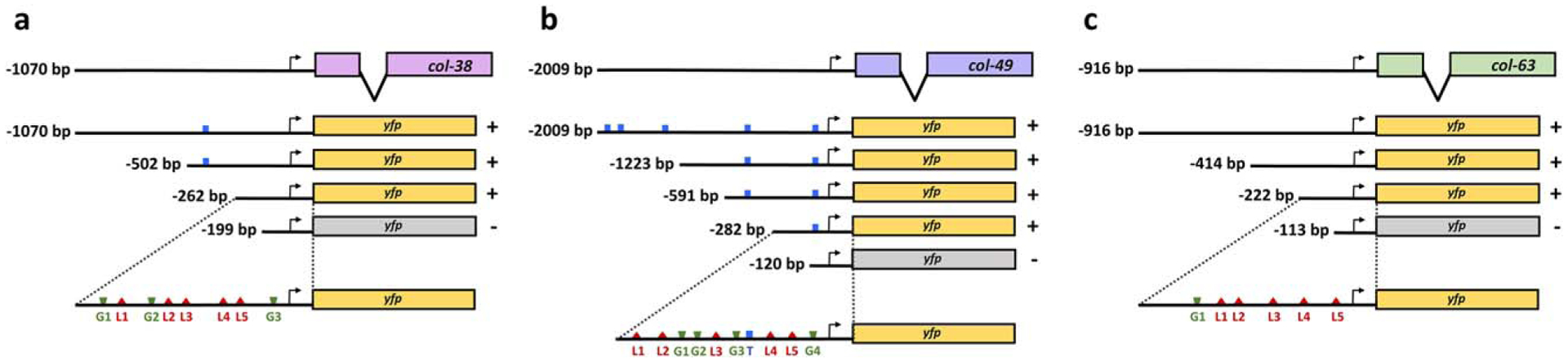
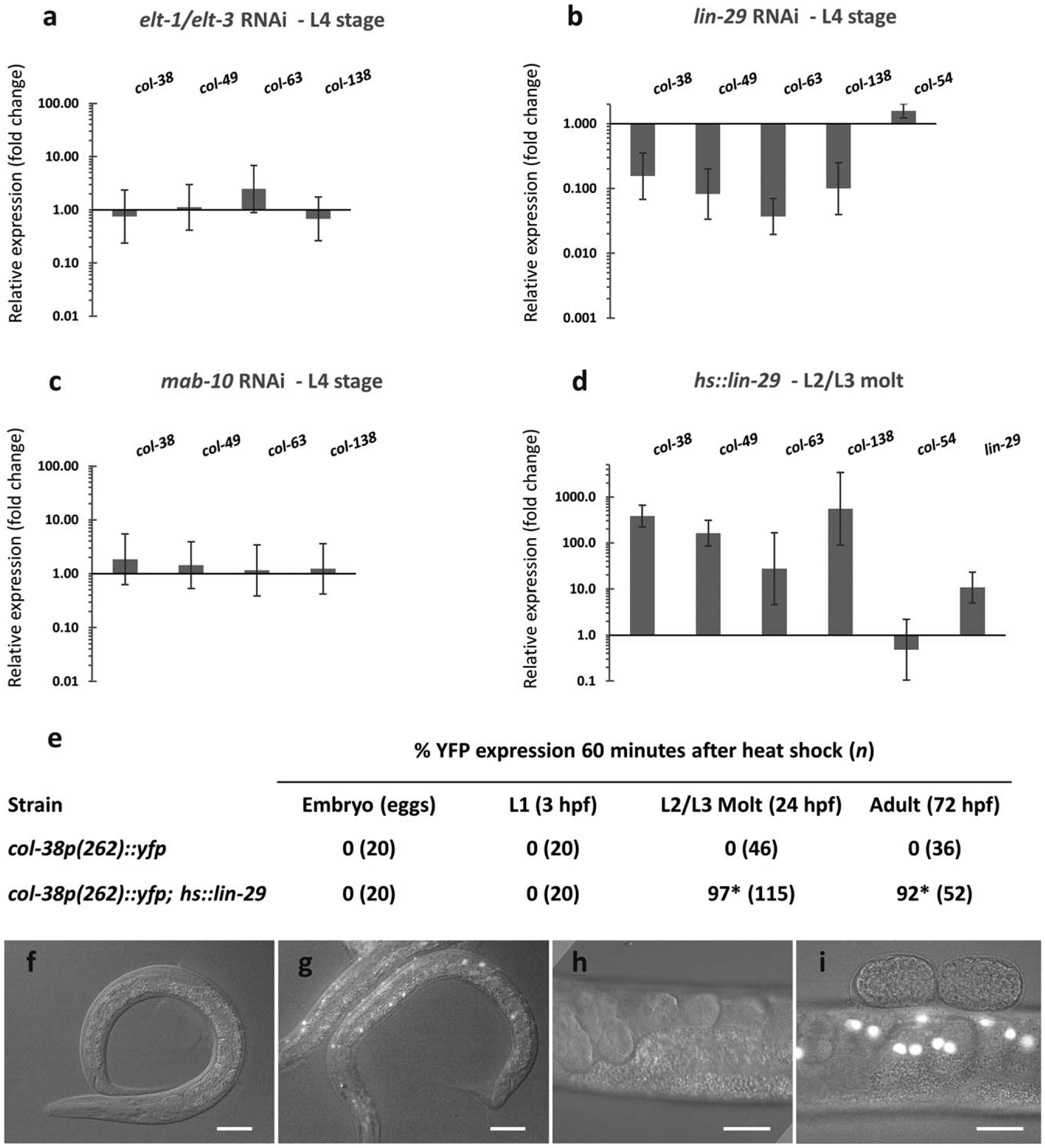
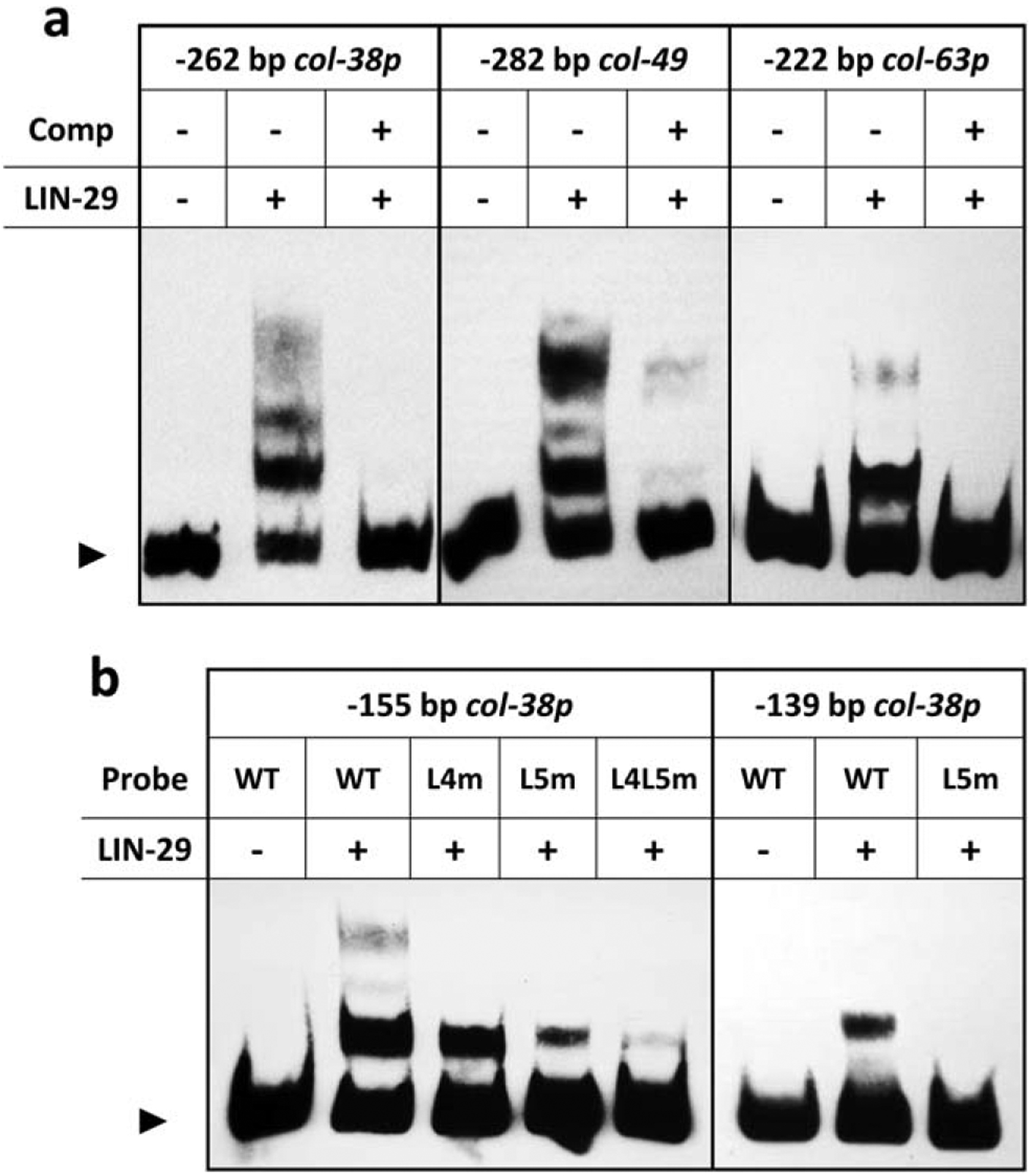
The cuticle, the outer covering of the nematode C. elegans, is synthesized five times during the worm's life by the underlying hypodermis. Cuticle collagens, the major cuticle component, are encoded by a large family of col genes and, interestingly, many of these genes express predominantly at a single developmental stage. This temporal preference motivated us to investigate the mechanisms underlying col gene expression and here we focus on a subset of col genes expressed in the L4 stage. We identified minimal promoter regions of <300 bp for col-38, col-49, and col-63. In these regions, we predicted cis-regulatory sequences and evaluated their function in vivo via mutagenesis of a col-38p::yfp reporter. We used RNAi to study the requirement for candidate transcription regulators ELT-1 and ELT-3, LIN-29, and the LIN-29 co-factor MAB-10, and found LIN-29 to be necessary for the expression of four L4-specific genes (col-38, col-49, col-63, and col-138). Temporal misexpression of LIN-29 was also sufficient to activate these genes at a different developmental stage. The LIN-29 DNA-binding domain bound the col-38, col-49, and col-63 minimal promoters in vitro. For col-38 we showed that the LIN-29 sites necessary for reporter expression in vivo are also bound in vitro: this is the first identification of specific binding sites for LIN-29 necessary for in vivo target gene expression.
Keywords: development; nematode; temporal expression.
© 2018 Wiley Periodicals, Inc.
Figures
References
-
- Brenner S (1974). The genetics of Caenorhabditis elegans. Genetics, 77, 71–94. Available at: http://www.ncbi.nlm.nih.gov/pubmed/4366476 - PMC - PubMed
-
- Cox GN (1992). Molecular and biochemical aspects of nematode collagens. Journal of Parasitology, 78, 1–15. Retrieved from http://www.jstor.org/stable/3283678 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
